Research Article - Der Pharma Chemica ( 2018) Volume 10, Issue 3
Comparison of Cellular Antioxidants, Enzyme Activities in Resistance and Susceptible Cucurbits Fruits Response to Melon Fly Infestation to Elucidate Possible Mechanism
Madhusudana Somegowda1,2*, Villangiri Vigneshwaran1, Shimoga V Sunil1, Shivashankar S3, Ramesh Babu HN4, Siddana Koppalu N Pramod1 and Achur N Rajeshwar2
1Department of Biochemistry, Sahyadri Science College (A), Kuvempu University Shimoga, Karnataka, India
2Department of Biochemistry, Janna Sahyadri, Shankaraghatta, Kuvempu University, Karnataka, India
3Department of Plant Physiology and Biochemistry, Indian Institute of Horticulture Research (IIHR), Hesaraghatta, Bangalore, Karnataka, India
4Department of Botany and Seed Technology, Sahyadri Science College (A), Kuvempu University Shimoga, Karnataka, India
- *Corresponding Author:
- Madhusudana Somegowda
Department of Biochemistry
Sahyadri Science College (A)
Kuvempu University Shimoga
Karnataka, India
Abstract
Melon fly is one of the serious pest worldwide infesting cucurbitaceous fruits inducing considerable damage resulting insufficient growth and economical loss. It is known that the susceptibility and resistance against melon fly infestation among cucurbittae fruits varies considerably. Henceforth, present study attempts to understand the cellular antioxidants potential and Phenyl Proponoid Pathway (PPP) enzyme expression among different cucurbittae fruits during infestation to compare susceptibility and resistance. This experiment conduct was healthy and melon fly infected cucurbittae fruits (Snake guard, bitter guard and bottle gourd) were selected and analyzed for phytoconstituents. In vitro antioxidant potential and expression of antioxidant enzymes and larvicidal activity in healthy and infected were compared. The PPP pathway enzyme level and expressions were also compared in healthy and infected fruit samples. Melon fly infestation considerably increases the contents of phenolic and flavanoids components in cucurbittae fruits compared to their healthy counterparts. Antioxidant enzymes expression found high in infected tissue samples as compared to healthy and apparent healthy. These observations indicate melon fly infestation induces cellular oxidative damage which was defended by increased antioxidant potential in infected fruits. The PPP pathway enzymes ((Phenylalanine Ammonia-Lyase (PAL) and Tyrosine Ammonia-Lyase (TAL), Cinnamyl Alcohol Dehydrogenase (CAD)), were also seen increased in expression by infected tissues which define the increased lignin synthesis at infestation site to provide resistance. Present comparative observations conclude bottle guard is more resistant whereas snake guard and bitter guard are susceptible for melon fly infestation. The resistance provided is linked with increased antioxidant potential and PPP pathway enzyme expression involved in lignin synthesis which oppose cellular damage and melon fly development and infection. Importance role of latex lectin is defense mechanism against insect, pathogens. There larvae activity of latex lectin has been importance documentation used as naturally occurs larvae with pest.
Keywords
Melons fly, Lignin, Phenol proponoid pathway, Bottle gourd, Larvae and latex
Introduction
Melon fly is one of the serious pest worldwide infesting cucurbitaceous fruits inducing considerable damage resulting insufficient growth and economical loss. The field experiments on assessment of loss of cucurbit fruit by melon fly infestation have been reported [1]. Depending on the environmental conditions and susceptibility of cucurbits species, the extent of losses varies between 30 to 100% [2]. Among all, bitter gourd (Momordica charantia L) is the worst affected with the rate of melon fly infestation as high as 95% and bottle guard is the least affected [3]. Plants have evolved sophisticated biochemical mechanisms to exert self-defense against pathogen, insect infection and UV-light protection [4]. The fruits of cucurbits consists biologically active secondary metabolites like phenols, flavanoids, terpenes, proteins, steroids, alkaloid and lignin, these help in defense mechanisms and increases the resistance against infection [5]. The increased expression of antioxidant enzymes during the infection is known to defend the potentially oxidative damage to the cells induced by the infected pests. The enzymes scavenge the reactive oxygen species and protect the cellular integrity by minimizing cellular destruction induced by invasion of pest [6]. Many studies have been showed the increased levels of secondary metabolite and antioxidant enzymes during pest infestation however, not much has been reported in regard to cucurbittae fruit resistance or susceptible during melon fly infestation.
The cucurbits fruits defense mechanism and Reacting Oxygen Species (ROS) produce to different stress and secondary metabolites signaling. ROS is widely reacting and induce damage to host tissues if not removed by scavenging mechanisms involving anti-oxidative enzyme [7]. The increased antioxidant enzymes in fruit develop resistance against pest induced damage to host. Many aspect of resistance is still unknown and the proper understanding to resistance fruits and differential immunity at biochemical and physiological level would help to devise strategies for control of the disease [8]. Numerous studies were also done to understand Phenyl Proponed Pathway (PPP) and its involvement in pest resistance by inducing lignin synthesis which harden the cell and provide protection by not allowing the pest to infect further to induce damage to cellular tissues [9]. In an extensive oviposition punctures induced by melon fly were seen on cucurbittae fruits for egg hatching and development of larvae in susceptible varieties however, surprisingly absent in resistance varieties [10], these observations report the low lignin content in susceptible fruits as compared to resistance fruits. It has been hypothesized that lignin synthesis during infestation might have a possible involvement as barrier for egg hatching and larval development of pest and play an important role as defensive component [11]. Plants have also evolved a variety of defense strategies to withstand the damage caused by herbivores. These defensive strategies may be expressed constitutively, or may be induced following herbivore infestation [12]. The resistance fruits are based on biochemical strategies required for the enzymes involved in PPP which necessities structural barrier against melon fly infestation along with secondary metabolites produced in response to infestation in cucurbits fruits defend against induced oxidative and biotic stresses [13]. With this present study attempts to compare the levels of metabolites, oxidative enzymes and PPP enzymes in susceptible and resistance variety of selected cucurbitae fruits after different stage of melon fly infestation. The varied levels may infer to identify the possible resistance mechanism offered to protect the fruit protection and destruction during infestation.
Material and Methods
Bitter gourd, Bottle gourd and Snake gourd seeds were obtained from January 14th 2012 in Horticulture department Shimoga, were cultivated in botanical garden of Sahyadri science college (A) Shimoga, Karnataka, India. The harvested fruits were infected by Oviposition of melon fly eggs. Eggs were the gifts from Entomology department, Indian Institute of Horticulture Research (IIHR), Bangalore. Most chemicals used during these experiments are obtained from Hi media and Merck, All the general chemicals and solvents of analytical grade.
Melons fly infestation on cucurbits fruits and assessment of infection
Freshly harvested bitter gourd, snake gourd and bottle guard matured fruits were selected for melon fly infestation. A circular hole measured 5 mm in diameter and 2 mm depth was punched on the surface of each fruit. The puncher spot was ovipositioned with 8-10 melon fly eggs. The eggs were allowed to hatch to produce melon fly larva. The larva fed on the fruit to increase in size to transform into pupa. The larval feeding induce the damage to fruit and was graded depend on the number of larval infection established and intensity of tissue damage.
The melon fly infested cucurbits were subjected to sampling of tissues as described previously [7] to assess the damage. The tissues were graded into Infected (A), Apparent Healthy (AH) and Healthy (H) group based on tissue damage and larval number in the infected spot.
Determination of lignin content in cucurbits fruits
Lignin content were followed by [13] cucurbits fruits healthy and infected fruits samples were extracted for 48 hr with methanol, and centrifuged for 5000 rpm for 15 min, collected pellet was dry in a incubator and ground to a powder, 500 mg of dry sample was mixed in 5 ml of 2 M HCl and 0.5 ml of thioglycolic acid, samples were heated at 95°C for 4 h and centrifuged at 3000 g for 20 min. Pellets were washed twice with de-ionized water, extracted successively with 2 ml of 0.5 M NaOH for 18 h and 2 ml of de-ionized water at room temperature and separated by centrifugation at 15 000 rpm for 10 min. Both the supernatants were collected, mixed, acidified with 1 ml of concentrated HCl and stored at 5°C overnight. The residue was dissolved in 10 ml of 0.5 M NaOH and centrifuged at 15 000 rpm. The absorbance of the clear supernatant was measured at 280 nm.
Estimation of total flavanoid and phenolic content in healthy and infected fruit tissues
The total phenolic content was assayed after extraction from melon fly infested tissue as. Briefly, 100 ul of tissue samples and standard solution (Galic acid at 100, 200, 300, 400 and 500 μg/ml) were added with 8.9 ml of distilled water and 1 ml of Folin-Ciocalteu’s phenol reagent. The mixture was shaken and after 5 min, 10 ml of 7% of Na2CO3 and 4.9 ml of distilled water was added to make a total volume of 25 ml. The mixture was incubated at room temperature for 90 min and absorbance was read at 750 nm [14]. Total phenolic acid content was expressed as μg/ml of Gallic Acid Equivalents (GAE) in the infected and healthy samples.
Total flavonoid content was measured by the aluminium chloride colorimetric assay [15]. An aliquot (1 ml) of extracts or standard solutions of quercetin (20, 40, 60, 80 and 100 μg/ml) was added to 10 ml volumetric flask containing 4 ml of distilled water. To the flask was added 0.30 ml 5% NaNO2, after 5 min 0.3 ml 10% AlCl3 was added. After 5 min, 2 ml M NaOH was added and the volume was made up to 10 ml with distilled water. The solution was mixed and absorbance was measured against the blank at 510 nm. The total flavonoid content was expressed as mg Quercetin Equivalents (QE).
Assay of non-enzyme antioxidant potential of cucurbittae fruit extracts
Cucurbittae fruit extracts from healthy and infected tissues were subjected non-enzymatic antioxidant assays following standard methods.
Ferric Reducing Antioxidant Power (FRAP)
FRAP assay was carried out following the published method [16]. Healthy and infected fruit sample (5 g) was homogenate with 100 ml of sodium acetate buffer (l M, pH 3.6), and filtered in muslin cloth, 100 μl of filtered sample was mixed with 3 ml FRAP reagent (Containing 2,4,6- Tripyridyl-S-Triazine (TPTZ), l M HCl and l M FeCl3 at 10:1:1 ratio) and tubes were vortexed and further incubated in boiling water bath for 30 min at room temperature and absorbance was read at 593 nm.
Super oxide onion (O2): Superoxide anion (O2) levels were determined by Nitroblue Tetrazolium (NBT) assay [17], 100 μl tissue homogenate was suspended in a solution containing 3 ml of Tris-HCl buffer (16 mM, pH 8.0), 1 ml of NBT (50 mM) and 1 ml Nicotinamide Adenine Dinucleotide (NADH) (78 mM), to that, add 1 ml of 10 mM Phenazinemetho-Sulphate (PMS) and incubate the mixture at 30°C for 10 min and read the absorbance at 560 nm.
2, 2-Diphenyl-1-Picrylhydrazyl (DPPH) free radical scavenging assay
Methanol extract of healthy and infected tissue was prepared [18]. In briefly, different concentrations of sample 20, 40, 60, 80 and 100 μl with mixed 1.5 ml of DPPH, mixture was shaken and allowed to stand at room temp (Dark condition) for 30 min. then, absorbance was measured at 517 nm.
Assay of antioxidant enzymes level and activity among infected and healthy cucurbits fruits
Different cucurbits fruit extracts with melon fly infected tissues were taken as used to antioxidant activity methods. Antioxidant enzymes level of extracted for taken 10 g of cucurbits healthy and infected tissue, were homogenate with 100 M phosphate buffer (pH 7.2). The homogenate was centrifuged at 10000 rpm for 15 min, at 10°C. The supernatant obtained was used for the determination to antioxidant enzymes activity.
Poly Phenol Oxides assay (PPO)
Polyphenol oxides activity were determine according to [19], take 10 g of cucurbits healthy tissue and melon fly infected tissue with homogenate 30 ml of 0.2 M phosphate buffer (pH 6.5) and 2 to 3 drop of Triton X-100 was added, were centrifuged at 10,000 rpm/30 min, then collected supernatant fruit sample used to further studies. The assay was contain in 3 ml of reaction miter, consist of 2.5 ml of (0.02 M pyrogallol and distilled water), with different concentration (20-100 μl/mg) of fruits sample. Reaction mixtures were incubated at 20°C for 30 min and absorbance at 420 nm were measured spectrophotometer.
Peroxidase (POD) assay
Peroxidase activity were analyzed by [20], 5 g of cucurbits fruit tissue with homogenate using chilled 4 ml of 100 mM sodium phosphate buffer (pH 7.0) and the filtered through four layers of muslin cloth and centrifuged at 18,000 rpm for 15 min at 4°C. The resulting supernatant are used as the crude enzyme source. Then 100 ul of cucurbits fruit tissue sample add 1.5 ml of phosphate buffer (pH 7.0) and 100 ul H2O2 with 1 ml of o-dianisidine solution and absorbance were recorded at 460 nm.
Superoxide Dismutase (SOD) activity
Cucurbits healthy and melon fly infected fruit tissue were, extraction with 10 ml of 65 mM potassium phosphate buffer (pH 7.8) and centrifuged at 10,000 g for 10 min. collected were supernatant, briefly take 100 μl of sample contained 50 mM sodium phosphate buffer, pH 7.8, 100 mM Ethylenediaminetetraacetic acid (EDTA), 130 mM methionine, 0.75 mM NBT, 20 μM riboflavin. The reaction was started by illuminating the tubes under four 40 W fluorescent lamps for 8 min, at the end of which time the absorbance at 560 nm [21].
Catalase (CAT) assay
Catalase activity is determination by [22], take healthy and infected different cucurbits tissue with homogenate in 100 mM potassium phosphate buffer (pH 7.0) and centrifuged at 18000 g for 15 min at 4°C, collected clear enzyme sample were used in the assay, content 100 mM potassium phosphate buffer (pH 7.2) and various fruit sample at 20-100 μl with 200 mM H2O2 added, absorbance at 240 nm.
Assay of Phenyl Proponoid Pathway (PPP) enzymes involved in lignin synthesis
Cucurbittae fruit extracts from healthy and melon fly infected tissues for PPP enzymes involved in lignin following methods.
Phenylalanine Ammonia-Lyase (PAL) and Tyrosine Ammonia-Lyase (TAL)
The enzymes activity using the method of [23], five gram of cucurbits fruit tissue were extracted in 10 ml of sodium borate buffer 50 mM, pH 8 and 8.5 for PAL and TAL, respectively. The homogenate was centrifuged at 5000 rpm for 15 min at 4°C, the supernatant used to enzymes assay. The reaction mixture consisting of 0.5 ml of enzyme extract and 150 mM of L-phenylalanine or L-tyrosine was adjusted to 3 ml with the extraction buffer. Incubation was done at 30°C for TAL and 40°C for PAL for 30 min. PAL activity was determined at 290 nm, following the formation of t-cinnamic acid and that of TAL at 310 nm by formation of p-coumaric acid.
Cinnamyl Alcohol Dehydrogenase (CAD)
CAD activity is based on [24], the reaction mixture in a contain volume of 3 ml contained 60 μl of 1 mM (E)-Cinnamaldehyde (Made up in 95% of ethanol), 600 μl of 1.0 mM Nicotinamide Adenine Dinucleotide (NADPH), 600 μl of enzyme extract, 50 μl of 20 mM ZnSO4.7H2O and 1150 μl of 0.1 M KH2PO4/Na2HPO4 buffer (pH 7.6) was added, and 15 min of incubation at 30°C was recorded spectrophotometrically at 340 nm.
Larval food medium and sample preparation
Second instar larval of melon fly were used for the larval activity, and maintained in an environment condition at 28°C. Medium for the larval growth and development was prepared by adding 20% sugar, 10% yeast dissolved in 100 ml of saline buffer and cucurbitaceae tissue sample, peeled skin and latex sap sample with concentrations of 1:1 and 1:5 ratio in μg/ml with medium, are taken for activity.
• Control-50 μl medium+200 μl of saline (Without sample).
• 1:1 sample-50 μl medium+100 μl saline+100 μl sample.
• 1:5 sample-50 μl food+160 μl saline+40 μl sample.
Larvicidal activities for melon fly
The different lectin sample of cucurbitaceae with medium was taken in small watch glass about 250 μl and second instar larva (Duplicate), were placed in it and are incubated for different time period with are observed for their weight viability and mortality.
Preparation crude protein sample
The crude protein sample was prepared from 10 mM Phosphate buffer saline (pH 7.2), different cucurbits fruits were taken and damaged fruit skin, after latex sample collect and were mixed in Phosphate-Buffered Saline (PBS) 1:9 (w/v) ratio. The mixed latex sample in centrifuged (REMI-R 8C DX) at 1000 × g for 10 min at 4°C and used further activity.
Haemagglutination (HA) activity
HA tests were done as per the methods described by [25]. Informed consent was obtained from all healthy human volunteers for obtaining peripheral venous blood in the age range of 20-30 years. Briefly, a 2% suspension of human blood was added to an equal volume of serially diluted protein solution in an agglutination plate, gently mixed and incubated at 37°C for 1 h to visualize the agglutination.
Latex proteins in midgut extraction from melon fly larvae
After feeding larvae dissection of the midgut were homogenation in 500 ul of ice cold extraction buffer (0.15 M NaCl. 50 ul mM TRIS-HCl, pH 11) in centrifuged 1300 g for 10 min at 4°C and supernatant was used for further to Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE). It was determination that incubation period from 5 to 80 h [26].
Statistics analysis
This experiment and results expressed as mean ± SEM and analysis of variance by A-Z Analysis of Various (ANNOVA) protocols, also significant (P < 0.05) measurements.
Results
Susceptibility to Bacteroceracucurbitae infestation varies across cucurbits species
The melon fruit fly, Bacteroceracucurbitae is reported to damage 81 host plants and it is a major pest of cucurbitaceous vegetable [27]. The ovipositioned bottle gourd, snake gourd and bitter gourd fruits were investigated and graded for the level of melon fly infestations. Our results, demonstrated that bottle gourd with higher level of B. cucurbittae resistance which was evident from the minimal tissue damage and negligible larval numbers. But snake gourd and bitter gourd depicted maximal tissue damage with 53.2% and 48.2% respectively with higher level of B. cucurbittae larval numbers in the infected site (Figure 1A-I). Examining the dissected fruit revealed high invasiveness of the melon fly larva in the susceptible fruit pulp compared to that of the resistant bottle gourd. The fruit of infected snake gourd and bitter gourd showed brownish and yellow discoloration with enormous reduction of the whole body (Figure 2A-H). It is very clear from our study results that snake gourd and bitter gourd show strong susceptibility to the melon fly infestations leading to the extensive fruit damage.

Figure 1: A-Bitter gourd uninfected, B- Apparent healthy fruit, C-Melon fly infestation tissue D-Snake gourd healthy, E-Infestation fruit, F-Completely melon fly infestation and G- Bottle gourd healthy, H-Melon fly infected site view and I-Not infestation
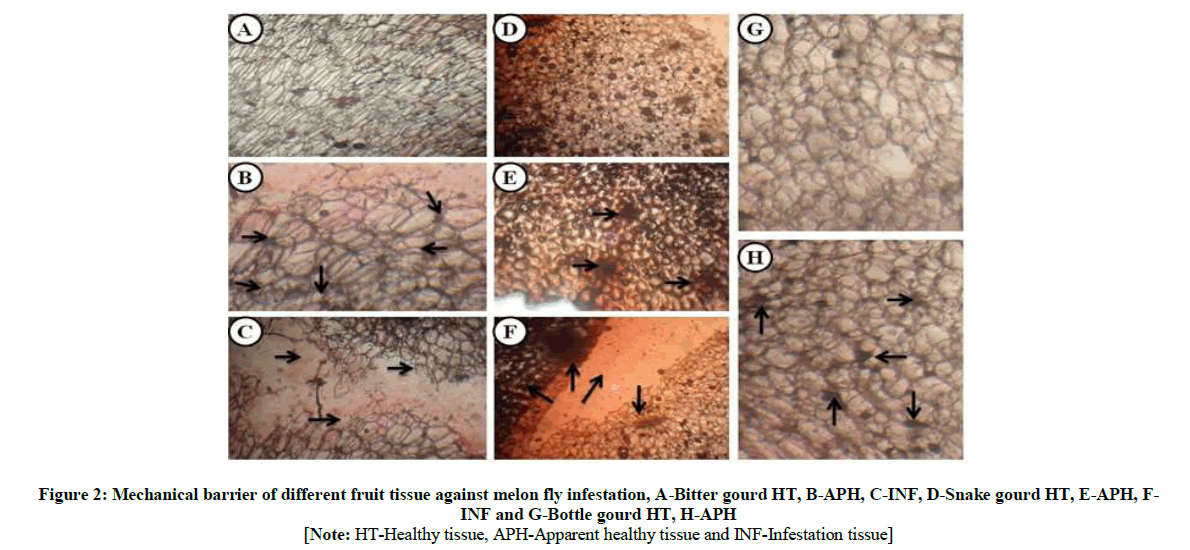
[Note: HT-Healthy tissue, APH-Apparent healthy tissue and INF-Infestation tissue]
Figure 2: Mechanical barrier of different fruit tissue against melon fly infestation, A-Bitter gourd HT, B-APH, C-INF, D-Snake gourd HT, E-APH, FINF and G-Bottle gourd HT, H-APH
Down regulated lignification corroborates with the B. cucurbittae susceptibility in cucurbits
Lignification is a mechanism for disease resistant in plants and is considered a first line defense against successful penetration of invasive pathogens [28]. In our study, B. cucurbittae infected tissues of Bottle gourd showed an increased level of lignin content whereas the bitter gourd and snake gourd showed lower levels of lignin deposition than the bottle gourd. Results clearly indicate that the lignin content in bottle gourd, snake gourd and bitter gourds were inversely proportional to the B. cucurbittae susceptibility (Figure 3). Moreover phenolic compounds and flavonoids are involved in host defense as a preformed chemical barrier that can prevent the pathogen penetration. Results elucidate that there was considerable increase in the total phenol and flavonoids in the bottle gourd leading to its potential resistance to the B. cucurbittae infestation (Figure 3A-C). But snake gourd and bitter gourd with negligible phenolic and flavonoid concentration showed higher susceptibility to the infection (Figure 3).
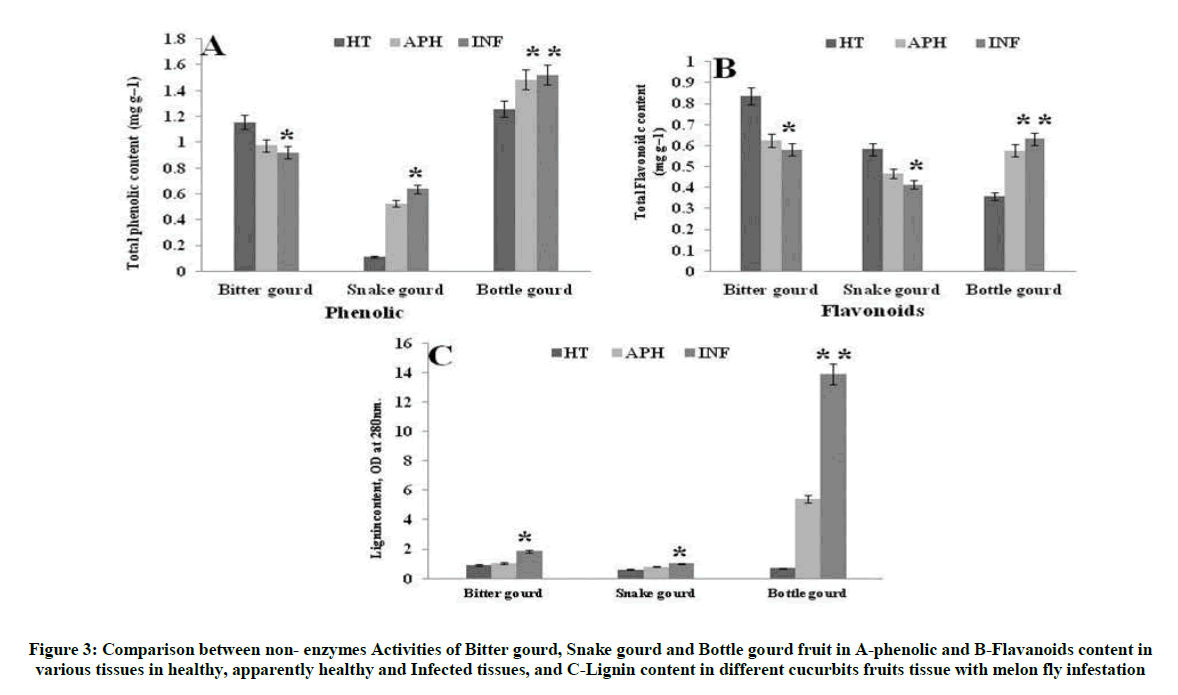
Figure 3: Comparison between non- enzymes Activities of Bitter gourd, Snake gourd and Bottle gourd fruit in A-phenolic and B-Flavanoids content in various tissues in healthy, apparently healthy and Infected tissues, and C-Lignin content in different cucurbits fruits tissue with melon fly infestation
B. cucurbittae infected cucurbits show severely declined enzymic antioxidants and ROS scavenging potential
Changes in activities of catalase, peroxidase and superoxide dismutase were investigated in the Healthy Tissue (HT), AH and Infected Tissue (INF) extracts of Bottle gourd, snake gourd and bitter gourd. The results elucidated that bottle gourd showed high levels of antioxidant enzymes in B. cucurbittae infected tissues compared to that of the INF of snake and bitter gourd. There was also a marked increase in the enzyme levels in the AH tissue of the bottle gourd as compared to the other two cucurbits. The results infer that the suppressed antioxidant enzyme levels in the snake gourd and bitter gourd resulted in increased susceptibility to the melon fly infestation. Supportively, declined enzymic antioxidants has reflected in the poor ROS scavenging potential of the INF and AH tissue of snake gourd and bitter as assessed by DPPH, hydrogen peroxide and superoxide scavenging assay. Whereas the INF and AH tissues of bottle gourd showed pronounced ROS detoxifying potential. These observations indicate there were declined antioxidant enzymes and ROS quenching activities in the susceptible snake gourd and bitter gourd, as compared to bottle gourd resistant B. cucurbitae infestation (Table 1).
| Parameter | Bitter gourd | Snake gourd | Bottle gourd | ||||||
|---|---|---|---|---|---|---|---|---|---|
| HT | APH | INF | HT | APH | INF | HT | APH | INF | |
| FRAP | 0.683 ± 0.15 | 0.524 ± 0.05 | 0.547 ± 0.48 | 0.621 ± 1.62 | 0.457 ± 0.17 | 0.410 ± 1.62 | 0.231 ± 0.82 | 0.275 ± 0.55 | 0.293 ± 0.08 |
| Super oxide anion (o2) | 0.053 ± 1.42 | 0.858 ± 0.01 | 0.921 ± 0.57 | 0.042 ± 0.21 | 0.638 ± 1.68 | 0.744 ± 0.13 | 0.086 ± 0.01 | 0.647 ± 0.28 | 0.718 ± 0.01 |
| DPPH | 0.673 ± 0.03 | 0.581 ± 1.96 | 0.314 ± 1.01 | 0.582 ± 0.03 | 0.428 ± 0.01 | 0.361 ± 0.02 | 0.003 ± 0.04 | 0.014 ± 0.01 | 0.017 ± 0.01 |
| PPO | 0.023 ± 0.01 | 0.029 ± 0.20 | 0.038 ± 1.05 | 0.044 ± 0.08 | 0.052 ± 0.01 | 0.064 ± 0.01 | 0.227 ± 0.38 | 0.251 ± 0.06 | 0.283 ± 0.61 |
| POX | 0.627 ± 0.68 | 1.870 ± 0.08 | 2.394 ± 0.01 | 1.242 ± 1.70 | 1.134 ± 0.01 | 1.142 ± 0.23 | 10.85 ± 0.45 | 12.34 ± 0.37 | 13.78 ± 0.32 |
| SOD | 8.412 ± 0.02 | 11.57 ± 0.02 | 17.63 ± 0.17 | 6.371 ± 0.22 | 10.52 ± 0.01 | 16.51 ± 0.07 | 7.156 ± 0.04 | 12.52 ± 0.01 | 19.44 ± 0.02 |
| CAT | 0.748 ± 1.81 | 1.689 ± 0.01 | 2.415 ± 1.04 | 0.372 ± 0.39 | 0.531 ± 0.05 | 0.968 ± 0.64 | 1.712 ± 0.04 | 3.62 ± 0.08 | 4.113 ± 0.08 |
Values are mean ± SEM, (n=1); *P < 0.05, **P < 0.001
Table 1: Comparison of Peroxidase activity as a measure of antioxidant potential in healthy, apparent healthy and infected tissue of Bitter gourd, Bottle gourd and Snake gourd against melon fly infection
Elevated PPP enzymes in cucurbits exerts defensive potential against B. cucurbitae
Phenylalanine Ammonia-Lyase (PAL) catalyses the first step in the biosynthesis of phenylpropanoids, which form a wide variety of defense related plant secondary products. Results infer that there was a remarkable up regulation of the PAL activity in the INF and AH tissues of bottle gourd. But INF tissues of snake gourd and bitter gourd demonstrated poor activity of PAL enzyme as compared to that of the bottle gourd. There was also severely lower levels of TAL and CAD in the INF and AH tissues of snake gourd and bitter gourd than the bottle gourd. The declined PAL, TAL, and CAD strongly correlates with the higher susceptibility to the melon fly infestation resulting in the increased B. cucurbitae larval deposition and fruit body decay (Figure 4A-C). Whereas bottle gourd accumulated defense related PPP enzymes in the infected site has lead to the minimal larval invasion and tissue damage. Therefore these results clearly indicate that possible role of PPP enzymes in providing resistance to the melon fly infestation in the cucurbitaceae vegetables.
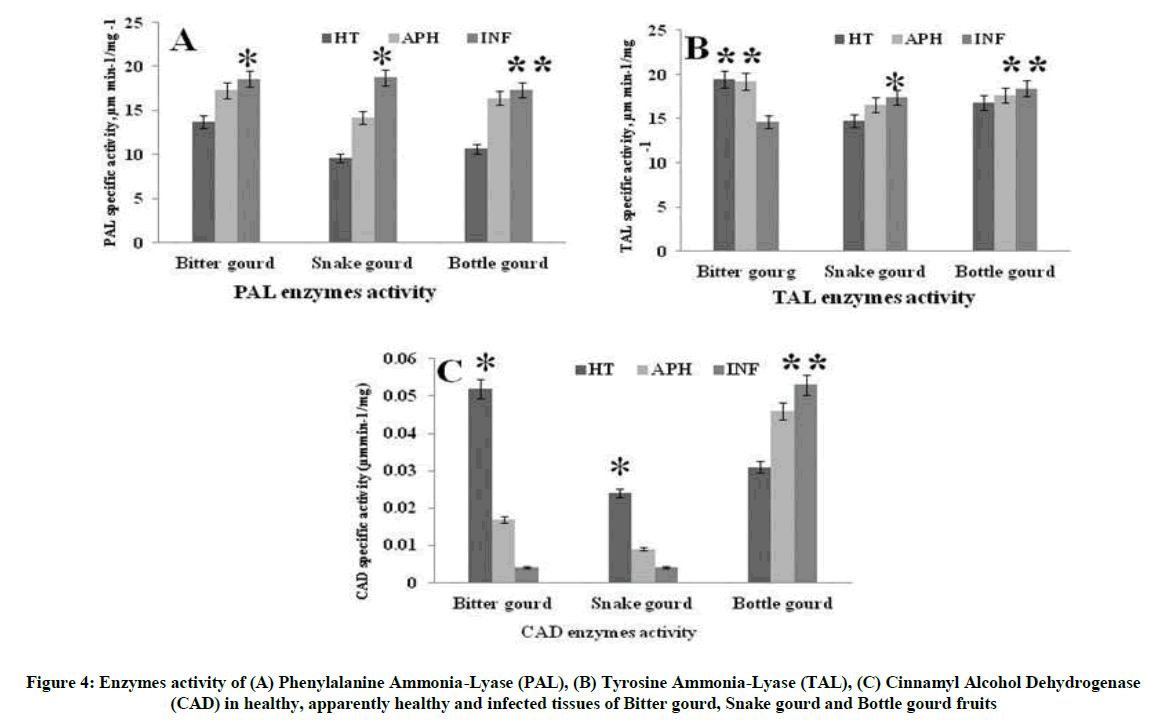
Figure 4: Enzymes activity of (A) Phenylalanine Ammonia-Lyase (PAL), (B) Tyrosine Ammonia-Lyase (TAL), (C) Cinnamyl Alcohol Dehydrogenase (CAD) in healthy, apparently healthy and infected tissues of Bitter gourd, Snake gourd and Bottle gourd fruits
Latex exudates of the resistant cultivar induces larval death in B. cucurbitae
The activity of second instar larvae was reduced by bottle gourd fruits latex sap more concentration (1:1) inhibits the mobility and weight reduced of the larvae compared to that of tissue samples and peeled skin samples of bottle gourd and snake gourd. Bottle gourd low concentration (1:5) also larvae activity and weight reduce, but take more time period compared to high concentration. Bitter gourd and snake gourd in various concentration and tissue sample did not show any larvae effect (Tables 2 and 3). Feeding latex sample to larvae some digestion problem in larval growth, also the discoloration of the skin of larvae is observed. Latex sample disturb to digestion problem and sticky like formation in midgut wall (Figure 5A-D). The agglutination assay is strongly inhibition to human irrespectively blood group show on (Figure 6A-C), partial purification latex lectin protein against the chick and human blood groups. Bottle gourd latex partial protein on the different numerical assay was strongly more stability.
| Time period | Bitter gourd | Snake gourd | Bottle gourd | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Skin | Latex | Tissue | Skin | Latex | Tissue | Skin | Latex | Tissue | ||||||||||
| 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | |
| 0-5 h | 0.014 g | 0.014 g | 0.013 g | 0.014 g | 0.013 g | 0.015 g | 0.013 g | 0.015 g | 0.014 g | 0.014 g | 0.015 g | 0.014 g | 0.014 g | 0.012 g | 0.013 g | 0.013 g | 0.015 g | 0.014 g |
| 5-10 h | 0.014 g | 0.014 g | 0.013 g | 0.014 g | 0.013 g | 0.015 g | 0.013 g | 0.015 g | 0.014 g | 0.015 g | 0.015 g | 0.015 g | 0.014 g | 0.012 g | 0.013 g | 0.014 g | 0.015 g | 0.014 g |
| 10-20 h | 0.015 g | 0.016 g | 0.014 g | 0.014 g | 0.014 g | 0.016 g | 0.014 g | 0.016 g | 0.015 g | 0.015 g | 0.016 g | 0.016 g | 0.015 g | 0.013 g | 0.014 g | 0.014 g | 0.016 g | 0.015 g |
| 20- 40 h | 0.016 g | 0.015 g | 0.014 g | 0.015 g | 0.015 g | 0.016 g | 0.015 g | 0.016 g | 0.015 g | 0.016 g | 0.017 g | 0.016 g | 0.015 g | 0.014 g | 0.013 g | 0.013 g | 0.016 g | 0.015 g |
| 40- 80 h | 0.017 g | 0.016 g | 0.015 g | 0.015 g | 0.016 g | 0.017 g | 0.016 g | 0.017 g | 0.016 g | 0.017 g | 0.017 g | 0.017 g | 0.014 g | 0.013 g | 0.013 g | 0.013 g | 0.017 g | 0.016 g |
Table 2: Effect of the cucurbits fruit Skin, Latex and Tissue in weight viability factor with second instar larvae for melon fly
| Concentration of various cucurbits fruits extract | Bitter Gour | Snake Gourd | Bottle Gourd | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Observation of Larvae activity | Skin | Latex | Tissue | Skin | Latex | Tissue | Skin | Latex | Tissue | ||||||||||||
| 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | 01:01 | 01:05 | ||||
| 0-5 h | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | |||
| 5-10 h | +++ | +++ | ++ | +++ | +++ | +++ | +++ | +++ | ++ | +++ | +++ | +++ | +++ | +++ | + | ++ | +++ | +++ | |||
| 10-20 h | ++ | ++ | ++ | ++ | +++ | +++ | ++ | +++ | ++ | ++ | +++ | +++ | ++ | +++ | --- | + | +++ | +++ | |||
| 20-40 h | ++ | ++ | ++ | ++ | +++ | +++ | ++ | ++ | + | ++ | +++ | +++ | --- | ++ | --- | --- | ++ | ++ | |||
| 40-80 h | ++ | ++ | + | ++ | +++ | +++ | ++ | ++ | + | ++ | +++ | +++ | --- | --- | --- | --- | ++ | ++ | |||
Table 3: Effect of the cucurbits fruit Skin, Latex and Tissue inhibition with second instar larvae Mobility for melon fly
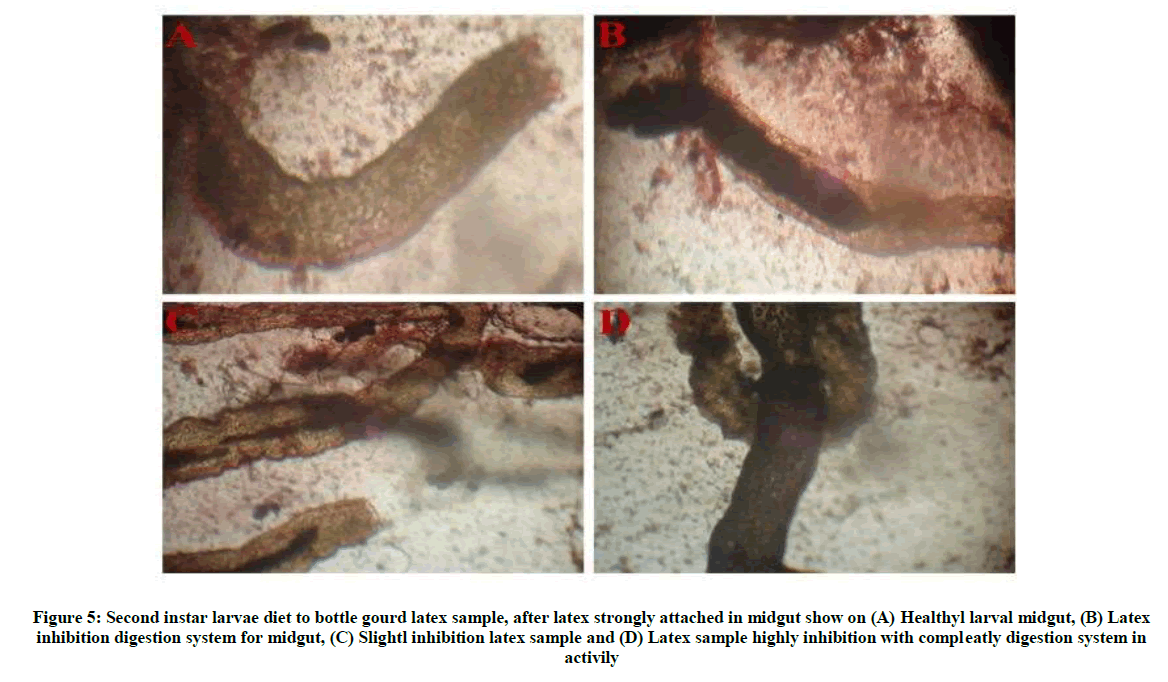
Figure 5: Second instar larvae diet to bottle gourd latex sample, after latex strongly attached in midgut show on (A) Healthyl larval midgut, (B) Latex inhibition digestion system for midgut, (C) Slightl inhibition latex sample and (D) Latex sample highly inhibition with compleatly digestion system in activily
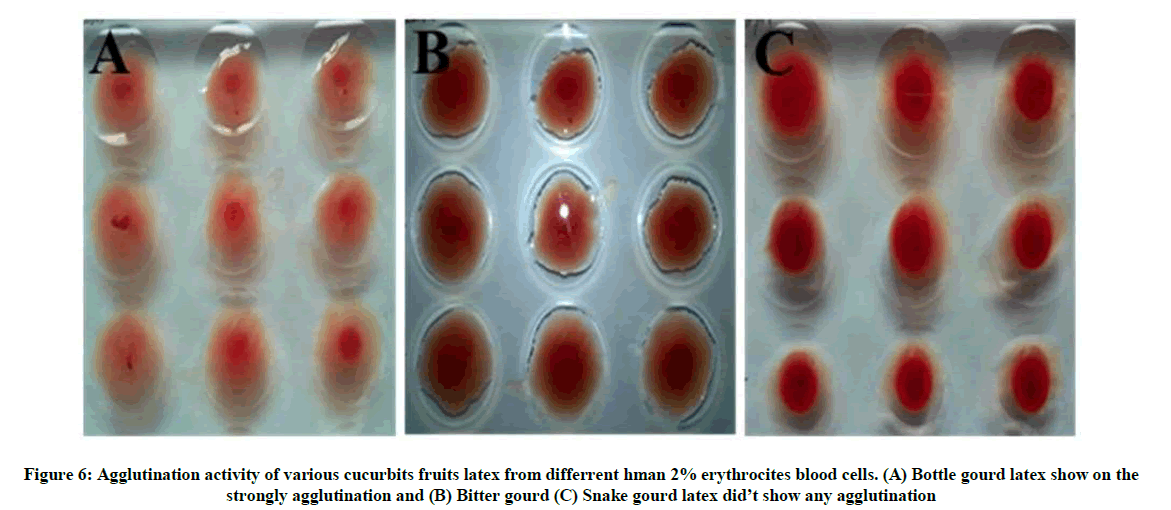
Figure 6: Agglutination activity of various cucurbits fruits latex from differrent hman 2% erythrocites blood cells. (A) Bottle gourd latex show on the strongly agglutination and (B) Bitter gourd (C) Snake gourd latex did’t show any agglutination
Discussion
The present studies of relation between cucurbits fruits with melon fly infestation, resistance fruit in can reduce Fe3+-TPTZ to its ferrous form; therefore, they possess higher potential antioxidant capacity [29] compared to susceptible fruits. Cucurbits healthy tissue in could not inhibits radical loses are inter mediated by ions but infestation fruit tissue not involved ions. The more variation level of superoxide radical in healthy fruits compared to melon fly infestation fruits. Melon fly infestation to resistance fruits of low level production of O2 radicals and moderately higher levels of melon fly infestation bitter gourd and snake gourd fruit. The significantly higher levels of O2 radicals generated in susceptible fruit compared to resistance fruit, in response to melon fly infestation, excessive levels of these free radicals have been implicated in the destruction of plant cells through the peroxidation of lipids and the generation of secondary cytotoxic species. Phenolic O-methylation and conversion of the side-chain carboxyl to an alcohol group [30] together with a substantial enrichment in free phenolic groups [31]. Melon fly infestation fruits in more endogenous level of phenolic and flavanoids contents will be synthesis, and important non-enzymatic antioxidants activity in host defense mechanism. Melon fly infestation cucurbits fruits in PPO activity Induces non- enzyme and enzyme scavenging activity in fruits and indicates their roles in host plant protection [32]. The PPO activity is more synthesized by various components such as quinine and inhibition of cellular enzymes, some protein, carbohydrates and altered components like dietary proteins, amino acids, flavanoids and phenolic also reduces their nutritional value to insect herbivores [33].
Lignin is a constituent of the cell walls of almost all fruits and plant cell wall, lignin to provide the structure and biological importance in fruits. Because it is much less hydrophilic than cellulose and hemi cellulose, it prevents the absorption of water by these polysaccharides in plant cell walls and allows the efficient transport of water in the vascular tissues [34]. Lignin also forms an effective barrier against attack by insects and fungi. Some resistance cucurbits fruits defense mechanism and protection to melon fly. POD scavenger activity is modification and development of different physiological stress in biosynthesis of some plant hormones and cell maintaining, cell adherence, cell breaking also resistance response. Activity rapidly increased in Bottle gourd infestation tissue compared to other than Apparent Healthy Tissue (APH) and HT, and are well scavenging activity and more lignin content producer. POD also promotes covalent linking between lignin and other cell wall polymers, providing a more effective barrier against insect attack [35], POD scavenging activity initiates toxic substances which are produced on the effected melon flies. Without SOD and enzyme designed to neutralize the superoxide anion, this molecule can cause immense damage at the cellular and molecular level. The extent of oxidative stress damage in leaf discs of both SOD+ and SOD- plants was dependent on their development [36], and very main function of SOD protection of cell against the toxic effects of superoxide radicals produced at different cellular loci [37]. SOD scavenging activity conversion of superoxide radicals to the more stable of H2O2 that is required for long-range cell-to-cell signaling or for passing through membranes [38-39]. The SOD activity increased in infestation tissue have been together more in H2O2 level and SOD activity level down in APH and HT. Considered to be a key player in the anti-oxidant defense system as it regulates the concentrations of superoxide radicals and H2O2, both of which have been shown to act directly or indirectly in plant defense and signal transduction [40]. CAT activity is maintaining the equilibrium reaction in physiological stress and under oxidative stress was found to be related to the accumulation of salicylic acid in plants [41], but CAT activity was dramatically decreased in plants under osmotic stress [42]. Salicylic acid is very importance roll resistance fruits in sending of biochemical signaling against melon fly insecticide, and CAT mutually helping plant physiological defense mechanism to ROS. Superoxide onion (O2) it is used by immune system to killing and protecting invading microorganism, insecticides and every time metabolism as by synthesis the needs for energy. Superoxide is highly toxic and from this one reactive oxygen species, numerous other toxic compounds can be generated such as the hydroxyl free-radical (OH), peroxynitrite (ONOO−) and H2O2.
PAL activity involving the elimination of amino group of phenylalanine to generate trans-cinnamate, which is converted to p-coumaric acid by the action of cinnamate 4-hydroxylase, while TAL activity leads directly to the production of p-coumaric acid from tyrosine [22]. Melon fly infestation Bottle gourd tissue was moderately increased PAL activity compared to Bitter gourd and Snake gourd, infestation tissue the synthesis of t- cinnamic acid rapidly increased in Bottle gourd. Increase of PAL in resistant line of cotton plant compared to the susceptible line after inoculation with Verticillum dahlia, TAL activity in increased was Bottle gourd infestation tissue compared to Bitter gourd and Snake gourd, TAL activity in p-coumaric acid gradually increased in bottle gourd infestation tissue. The t- cinnamic acid and p-coumaric acid highly produced in resistance fruits compared other fruits, it may be metabolic process will be change and p-coumaric acid was capability protection from microorganism, p-Coumaric acid is also the precursor of flavanoids which are related to defense responses in plants. CAD induction has been studied in relatively few plant defense responses, CAD reaction was PPP is leading to synthesis in lignin content and also additional of water molecules reaction. Determinations of lignin content in cucurbits fruits involved PPP with melon fly infestation. Finding on the rapidly increased in lignin monomer in resistance and decline susceptible fruits infestation have strongly indicated, the main roll of cellular antioxidant, non-enzymes and enzymes of pp pathway of lignin content in the resistance cucurbits fruits against infestation activity. Resistance fruits was found to be effective neutralization of free radicals while such a balancing mechanism and antibiosis targeting larval development and lack of damage, was absent in susceptible fruits. Melon fly attempt some time to resistance fruits attacked, reacting oxygen species synthesis and neutralization by anti-oxidants help to regain homeostasis, more level of POD in the latex of resistance and possibly, the involvement of sugars in signaling reactions related to host defense and PPP enzymes generating phenolic and flavanoids content and leading to extensive lignifications. On the contrary, susceptibility fruits are related to the weak and delayed response with respect to the generation of superoxide and H2O2 associated with the generation of disproportionately high level of hydroxyl radical and followed by ineffective removal by antioxidants [8].
The melon fly larvae growth activity is reported on reduced in larvae, food congaing bottle gourd latex highly binding to midgut, because larvae growth factor will be reduced also abnormal activity and not survival. Bottle latex sample diet to second instars larvae slightly abnormal activity affected show on the reported, and tissue sample diet larvae doesn’t affected and other both fruits various sample did not any larvae effect an growth viability and inhibition activity. The bottle gourd latex upon wounding that harbors more lectin protein properties and diverse metabolites in multifunctional therapeutic capabilities. Lectin is a diverse group of proteins that are able to specifically recognize cell-surface carbohydrates, thus playing central roles in biological processes and cellular signaling. The ability to agglutinate is a simple measure to the detect presence of lectin proteins [25]. Bottle gourd showed potent agglutination in both erythrocytes has strongly inhibition the presence of lectin, but did not any agglutination activity other Bitter gourd and snake gourd, now conformed bottle gourd is a resistance fruits. This study has identified that role of ROS, antioxidant, enzymes, proteins and secondary in the resistance fruit of host against fly infestation and bottle gourd play importance role in resistance with insect (Figure 7).
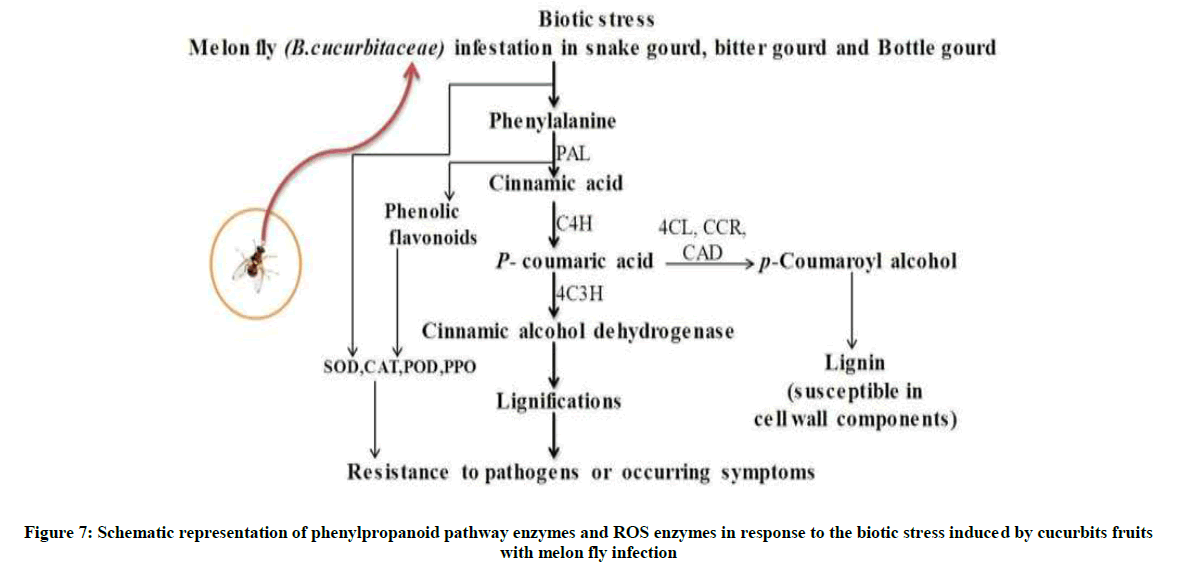
Figure 7: Schematic representation of phenylpropanoid pathway enzymes and ROS enzymes in response to the biotic stress induced by cucurbits fruits with melon fly infection
Acknowledgement
The authors kindly acknowledge the institutions, Indian Institute of Horticulture Research (IIHR), Bengaluru, and Kuvempu University, Shimoga for supporting this study. The authors are grateful to the Department of Entomology, IIHR for their kindness in providing Bacterocera cucurbitae pupa for conducting this study.
References
- P. Bwonmik, Indian J. Plant Prot., 2014, 1, 34-37.
- D. Gupta, A.K. Verma, Adv. Plant Sci., 1992, 5, 518-523.
- S.V. Singh, A. Mishra, R.S. Bisan, Y.P. Malik, A. Mishra, Indian J. Entomol., 2000, 62, 242-246.
- W.U. Cusui, Plant Physiol., 1997, 115, 427-435.
- S. Tanuja, Engineering Science and Technology: An Inter J., 2012, 2, 861-874.
- W. Mengyi, Front Plant., 2015, 6, 1-11.
- S. Shivashankar, J. Horticultural Sci. & Biotech.,2012, 87, 391-397.
- P. Shalini, Biomed. Informat., 2013, 20, 1013-1018.
- R.L. Johu, Walker, Biotech. Gentienge., 1998, 15, 1-42.
- M. Gavigowda, Submitted to University of Agricultural Sciences, Bangalore, 1976, 115.
- A. Gaylord, PANS., 2011, 108, 1-5.
- A. Kessler, I.T. Baldwin, Science., 2001, 291, 2141-2144.
- M.A. Doster, R.M. Bostock, American Phytopathological Society., 1988, 78, 473-477.
- A.A. Alafiatayo, Complement. Altern. Med., 2014, 11(3), 7-13.
- D. Patricia, Chinese Med.,2012,7, 26.
- A. Popova, D. Mihaylova, I, Alexieva, Int. J. Curr. Microbiol. App. Sci., 2014, 3(11), 436-444.
- N. Doke, Physiol. Plant Pathol., 1983, 23, 345-357.
- W. Aneta, Food Chem., 2007, 105, 940-949.
- K. Jae-Joon, k. Woo-Yeon, J. Ginseng Res., 2013, 37, 117-123.
- Ila Bania, Inte.r J. Scientific and Research Publications., 2012, 2250-3153.
- A.N. Yuyan, PLoS. One., 2016, 10, 1371.
- S. Shivashankar, Ann. Appl. Biol., 2015, 1-15.
- L.C. Cynthia, J. Experimen. Botany., 2015, 66, 4317-4335.
- J. Heidi, Mitchell, Plant Physiol., 1994, 104, 551-556.
- V. Vigneshwara, T. Prabhu, S. Madhusudana V. Krishna, S.N. Pramod, B.T. Prabhakar, Int. Immunopharmacol., 2016, 39, 158-171.
- V. Gianni, J. Experimen. Botany., 2010, 61, 1003-1014.
- M.K. Dhillon, R. Singh, J.S. Naresh, H.C. Sharma, J. Insect Sci., 2005, 1536-2442.
- H. Nazmul Bhuiyan, G. Selvaraj, Y. Wei, J. King, Plant Signaling & Behavior., 2009, 4(2), 158-159.
- M.M. Bruno, A.C. Pieme, P.C.N. Biapa, J.R. Njimou, M. Stoller, M. Bravi, Jeanne Y. Ngogang, Evid. Based Complement. Alternat. Med., 2015, 14.
- J. MacKay, T. Presnell, H. Jameel, H. Taneda, D. O’Malley, R. Sederoff, Holzforschung., 1999, 53, 403-410.
- F.E. Laveda, N. Delicado, F. Garcia-Carmona, A. Sanchez Ferrer, J. Agric. Food Chem., 2001, 49, 1003-1008.
- G.W. Felton, K.K. Donato, R.M. Broadway, S.S. Duffey, J. Insect Physiol., 1992, 38, 277-285.
- L. Ingham, M.L. Parker, K.W. Waldron, Physiologia Plantarum., 1998, 102, 93-100.
- S.A. Gupta, S.G. Ashima, Plant Physiol., 1993, 103, 1067-1073.
- H. Halliwell, B.J.M.C. Gutteridge, Plant Physiol., 2006, 141(2), 312-322.
- A.C. Allan, R. Fluhr, Plant Cell., 1997, 9, 1559-1572.
- E. Vranova, D. Inze, F. Breusegem, J. Experimen. Botany., 2002, 53, 1227-1236.
- R. Fabio, New Phytologist.,2004, 163, 563-571.
- L.L. Xu, L. Tu, L. Liu, D. Yuan, L.L. Jin, L.L. Long, X. Zhang, J. Experimen. Botany., 2011, 62, 5607-5621.
- I.S. Shim, Y. Momose, A. Yamamoto, D.W. Kim, K. Usui, Plant Growth Regul., 2003, 39, 285-292.
- H.A. Stafford, Flavonoid Metabolism, 360. Boca Raton, FL, USA: CRC Press, Boca Reton, FC, 1990.
- B.H. Moerschbacher, K.H. Kogel, O. Obst, H.J. Reisener, Z. Naturforsch., 1986, 41, 839-844.