Research Article - Der Pharma Chemica ( 2018) Volume 10, Issue 4
5-Amino Salicylic Acid and Sodium Salicylate Inhibit Activation of IκB Kinase and P38 γ MAP Kinase in LPS Induced RAW 264.7 cell
Saurabh Gupta1*, Gurjeet Thakur1, Sandeep Arora1, Rama Satyanarayana Raju Kalidhindi2, Nilesh S Ambhore2, Kannan Elango2, Ashish D Wadhwani3 and Md. Afzal Azam4
1Department of Pharmacology, Chitkara College of Pharmacy, Chitkara University, Chindigarh-Patiala National Highway, Rajpura, Panjab-140401, India
2Department of Pharmacology, JSS College of Pharmacy (Off campus JSS University), Udhagamandalam, The Nilgiris, Tamil Nadu-643001, India
3Department of Pharmaceutical Biotechnology, JSS College of Pharmacy, (Off campus JSS University), Udhagamandalam, The Nilgiris, Tamil Nadu-643001, India
4Department of Pharmaceutical Chemistry, JSS College of Pharmacy (Off campus JSS University), Udhagamandalam, The Nilgiris, Tamil Nadu-643001, India
- *Corresponding Author:
- Saurabh Gupta
Department of Pharmacology
Chitkara College of Pharmacy
Chitkara University
Chindigarh-Patiala National Highway, Rajpura, Panjab-140401, India
Abstract
Allergic asthma is a complicated disorder where multiple stimuli activate various signalling pathways resulting in the synthesis of a wide array of inflammatory proteins. Among the diverse transcription factors in the human genome, Nucleus Factor Kappa B (NFκB) and p38 Mitogen Activated Protein (MAP) kinases play a crucial role in the pathophysiology of allergic asthma. Salicylic acid derivatives, 5-Amino Salicylic Acid (5-ASA) and Sodium Salicylate (SS) have been evidenced to inhibit NFκB in vitro model. 5-ASA has demonstrated significant antiasthmatic activity in preclinical murine models. In the present study, 5-ASA and SS were screened in silico against a definite set of proteins, namely Inhibitor of Kappa Beta (IκB), p38 α, β, γ and δ using Glide and Prime modules of Schrodinger suite. The two drug candidates were attributed to the MTT assay in RAW 264.7 mouse monocyte macrophage cells for their in vitro cytotoxicity. From the hits obtained in silico, IκB and p38γ were further evaluated using western blot analysis followed by evaluation of TNF-alpha, IL-6, IL-13, nitrate and nitrite, Myeloperoxidase (MPO) and Malondialdehyde (MDA) levels in Lipopolysaccharide (LPS) induced RAW 264.7 cells. The results revealed significant reduction in the levels of the aforementioned parameters accompanied by the reduced expression of IκB and p38γ in cells treated with 5 ASA and SS, when compared LPS induced cells indicating the immune suppressive potential of both the drugs that can be used against allergic asthma.
Keywords
5-Amino salicylic acid, Sodium salicylate, NFΚB, P38 Map kinases, RAW 264.7 cells, Allergic asthma
Introduction
Transcription factors play a pivotal role in deciding the fate of a cell controlling all aspects in its regular functioning. A multitude of transcription factors and their role in cell functioning have been identified till date, few of which include Nucleus Factor Kappa B (NFκB), Map Kinases (P38 α, β, γ and δ), activator proteins, STATs and C-JNK which play a crucial role in regulating the immune responses of a cell [1]. NFκB and P-38 map kinases have gained significant prominence as targets for immune regulation in the scientific community.
NFκB is a pleiotropic transcription factor involved in the synthesis of TH2 cytokines, chemokines, adhesion molecules and inflammatory enzymes activated by TNF-α, oxidative stress and environmental toxins [2]. It is located in the cytosol as a heterodimer bound to the Inhibitor of Kappa B (IκB) and is activated by Inhibitor of Kappa B kinase which phosphorylates IκB leading to its activation and subsequent nuclear translocation [3]. The behavior of a cell depends on the intracellular signalling proteins such as MAP kinases [4]. MAP kinases are members of crucial signalling cascades (Extracellular signal-Regulated Kinases-ERK, AP signalling etc.) which serve as critical focal points in response to extracellular stimuli [5]. The MAP kinase family constitutes of four different components; extracellular signal-regulated kinases (ERKs), c-jun N-terminal or Stress-Activated Protein Kinases (JNK/SAPK), ERK/big MAP kinase 1 (BMK1) and the p38 group of proteins. Of these four, P38 MAP kinases are known to be responsible for aggravating immune responses in inflammatory disorders [6-8]. P38 MAP kinases constitute of 4 sub-groups like α, β, γ and δ and are categorized by a Thr-Gly-Tyr (TGY) dual phosphorylation motif, of which α and β are ubiquitously expressed in all cell types, while γ and δ are expressed in limited cell variants [9-14]. 5-ASA and SS have been proven to inhibit IκBK thereby inhibiting NFκB activation [15,16]. 5-ASA is also effective against a preclinical model of allergic asthma and is known to regulate the levels of TH2 cytokines and oxidative stress [17]; however, the exact mechanism of action remains ambiguous as to whether IκBK is being inhibited directly or by a simultaneous inhibition of a neighboring protein in the signaling cascade.
In this preliminary study we hypothesized and execute the attempt to screen 5 ASA and SS on IκBK and P38 MAP kinases via in silico followed by in vitro evaluation of protein expression and biochemical evaluation in LPS induced RAW 264.7 cell lines thereby providing a plausible explanation of the exact mechanism of action of the two drug candidates.
Materials and Methods
Chemicals and kits
NFκB primary antibody & P-38γ primary anti-mouse antibody were procured from Santa Cruz Biotech, USA. HRP conjugated antimouse IgG secondary antibody, Tumor Necrosis Factor-α (TNF-α) (Cat#: K0331186) and interleukin-6 (IL-6) (Cat#: K0331230) kits were obtained from Koma Biotech, South Korea. Interleukin - 13 kit (Cat#: ELM-IL13-001) was obtained from Ray Bio, USA. O-dianisidine, Flavin Adenine Dinucleotide (FAD), may-grunwald, Hexadecyltrimethylammonium Bromide (HTAB), thiobarbituric acid, N-[2-Hydroxyethyl]piperazine-N′- [2-ethanesulphonic acid] (HEPES), griess reagent, 1,1,3,3-tetraethoxypropane, reduced Nicotinamide Adenine Dinucleotide Phosphate (NADPH), nitrate reductase, Sodium salicylate, lipopolysaccharide, thiobarbituric acid, 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide (MTT), trichloroacetic acid, were purchased from sigma Pvt. Ltd. etc. Folin Ciocalteu's reagent was obtained from Loba Chemie, India. All chemicals used were of analytical grade. 5 ASA and Dexamethasone was obtained as a gift sample from Zydus Cadila and Ranbaxy Pvt. Ltd.
In silico screening
Ligand preparation
The structures of the ligands were generated in the CDX format using Chem Draw ultra-version 8.0. These ligands were subsequently converted to the mol format and optimized using Lig, Version 3.1. The 3D conversion and minimization was performed using optimized potential for liquid simulations (OPLS-2005) force field [18].
Docking
The X-Ray crystallographic structures of the proteins were retrieved from the Protein Data Bank: PDB ID: 4KIK [19] and PDB ID: 1CM8 [20] and were optimized using the Protein Preparation Wizard, [21] Schrodinger Suite 2014-3. Hydrogen atoms and charges were added using the OPLS 2005 force field and hydrogens were refined using restrained minimization (Figure 1). Prime version 3.7 was utilized for side-chain refinement and to build breaks present in the structure. Ligands were desalted and tautomers were generated. OPLS-2005 force field was used for the geometric optimization. Generation of multiple ligand ionization states and tautomeric forms at pH 7.0 ± 0.2 was performed using Epik, version 2.9 of Schrodinger suite [22]. Additional preparations of protein and ligand were performed using default parameters. The ligand-receptor interactions and dock poses of the ligands are of importance to quantify the strength of association of ligands with receptor [23]. The docking study into the catalytic domain of 1CM8 and 4KIK was performed for both the ligands in flexible mode using Glide, version 6.4, Schrodinger 2014-3 [24] by enabling the “write XP descriptor information” and calculate the docking scores.
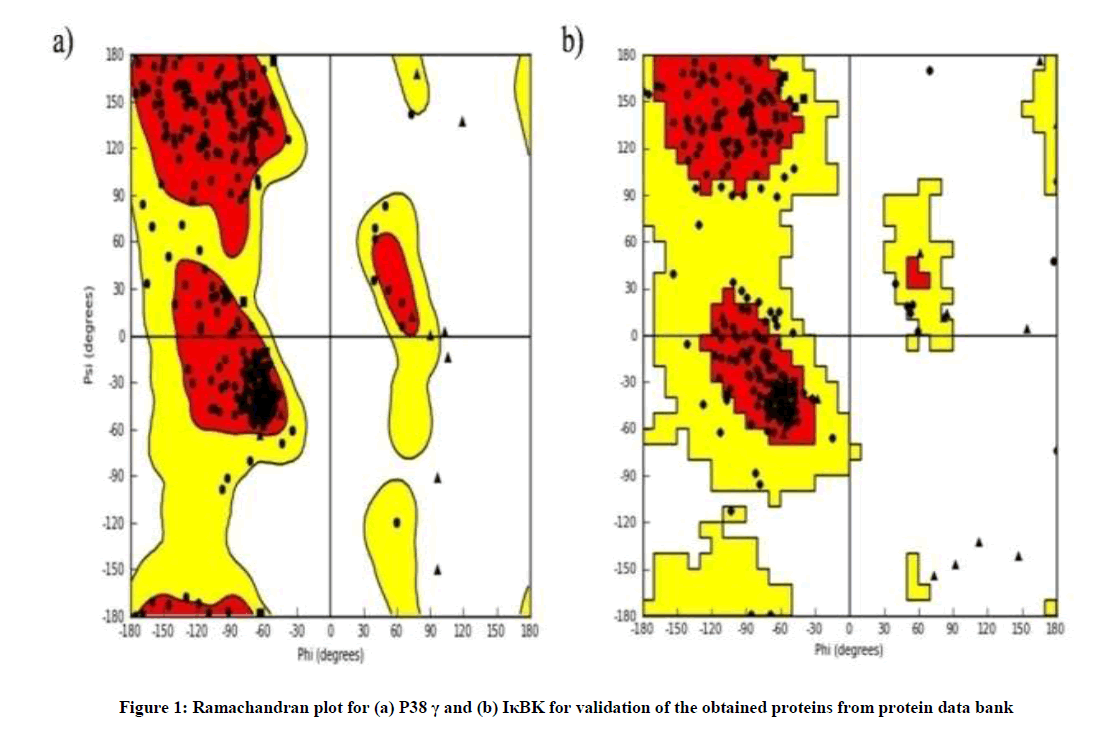
Figure 1: Ramachandran plot for (a) P38 γ and (b) IκBK for validation of the obtained proteins from protein data bank
In vitro cytotoxicity
MTT assay was performed for 5 ASA and SS in addition to standard dexamethasone in RAW 264.7 cells for cytotoxicity. Each 96 well plate was seeded with 8000 cells and incubated at 37°C for 24 h prior to initiation of the assay. The seeded wells were treated with respective drug solutions at test doses (10, 20 and 30 μg/ml) followed by addition of MTT (10 μl) after 4 hours [25]. The absorbance was recorded at 570 nm (Table 2).
In vitro screening
Raw 264.7 cells were cultured in Delbecco’s Modified Eagle’s Medium (DMEM) + 10% Foetal Bovine Serum (FBS) as per standard procedure. 5 × 106 cells were taken and incubated in a 24 well plate. All the cells except for normal control were activated using LPS at a concentration of 1 μg/ml for 15 min [26]. The cells were treated at three concentrations of test samples (10, 20 and 30 μg/ml), a normal control, negative control (LPS induced) and standard (Dexamethasone).
Apoptosis study
RAW 264.7 cells were digested with 0.25% trypsin (Hyclone, Logan, UT). DMEM with 10% FBS was deposited in each well of a 96-well plate (100 μl/well). Cells were added to a final concentration of 2 × 10/ml and the plates were incubated. Cells were left untreated or treated with 10, 20, or 30 μg/ml of 5-ASA and SS. The samples in a 96-well plate were divided into 4 groups, with 24 well samples in each group corresponding to different reagent concentrations. After being cultured for 24 h, 20 μl of trypsin was added into each well. When cells had sloughed off, suspensions (25 μl) were transferred to glass slides. Dual fluorescent staining solution (1 μl) containing 100 μg/ml Acridine Orange (AO) and 100 μg/ml ethidium bromide (EB) was added to each suspension and then covered with a cover slip. The morphology of apoptotic cells was examined and 500 cells were counted within 20 min using a fluorescent microscope (OLYMPUS, Japan) [27]. Dual AO/EB staining method was repeated 3 times at least.
Western blot
The monocyte-macrophage cells subjected to the above mentioned treatment were subjected to western blot analysis. The cell suspension was centrifuged at 3000 rpm for 10 min to obtain a pellet. The pellet was re-suspended in three volumes of lysis buffer (100 mM/l KCl, 10 mM/l HEPES, 1 mM/l EDTA, 1% Triton X-100 and protease inhibitor). The suspension was centrifuged at 2000 rpm for 10 min and the supernatant was subsequently centrifuged at 10,000 rpm for 30 min at 4°C. The final pellets were re-suspended in 100 μl of lysis buffer and mixed with the supernatant obtained after second centrifugation. The total protein concentration was measured by Bradford’s method [28]. 50 μg of protein from each sample was subjected to separation by Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) using 8% polyacrylamide gels followed by transfer of proteins to a nitrocellulose membrane. The membranes were then incubated overnight with mouse anti IκBK and P-38γ (1:1000) respectively at 4°C. Anti-mouse IgG-HRP (1:2000) was added at room temperature and incubated for 60 min as a secondary antibody. Quantification was performed using densities of each sample band. The images were developed using an image scanner connected to a computer.
Expression of TNF-α, IL-6 and IL-13
The levels of TNF-α, IL-6 and IL-13 were determined using Enzyme-linked Immunosorbent Assay (ELISA) kits according to the manufacturer’s protocol.
Estimation of nitrate and nitrite levels, myeloperoxidase, malondialdehyde
The levels of nitrate and nitrite were estimated according to the method reported by Toward et al., [29]. Malondialdehyde activity was determined using the method specified by Aruoma et al., [30]. Measurement of myeloperoxidase activity in lungs was performed by the method specified by Bradley et al., [31].
Statistical analysis
Statistical analysis was carried out by using One-Way Analysis of Variance (ANOVA) followed by Tukey's multiple comparison tests. P-value b 0.05 was considered statistically significant. The analysis was carried out using GraphPad Prism software V.5.04.
Results
Binding potential of 5-ASA and SS with IκBK and P38 γ proteins in silico
The results reveal interactions of 5 ASA with P-38γ and IκBK through hydrogen bonding with amino acids LYS 56 with a glide score of -8.60 and GLU 97 with a glide score of -6.40 respectively. Similarly, SS revealed hydrogen bonding with P 38γ at GLY 157 and IκBK at GLU 97 and LYS 99 with glide scores of -8.33 and -6.15 respectively (Figure 2). In case of the standard, dexamethasone interacted with p38 γ at GLY 157 with a glide score of -8.40 while IκBK at ASP 166 and THR 23 with a glide score of -6.20 respectively (Table 1).
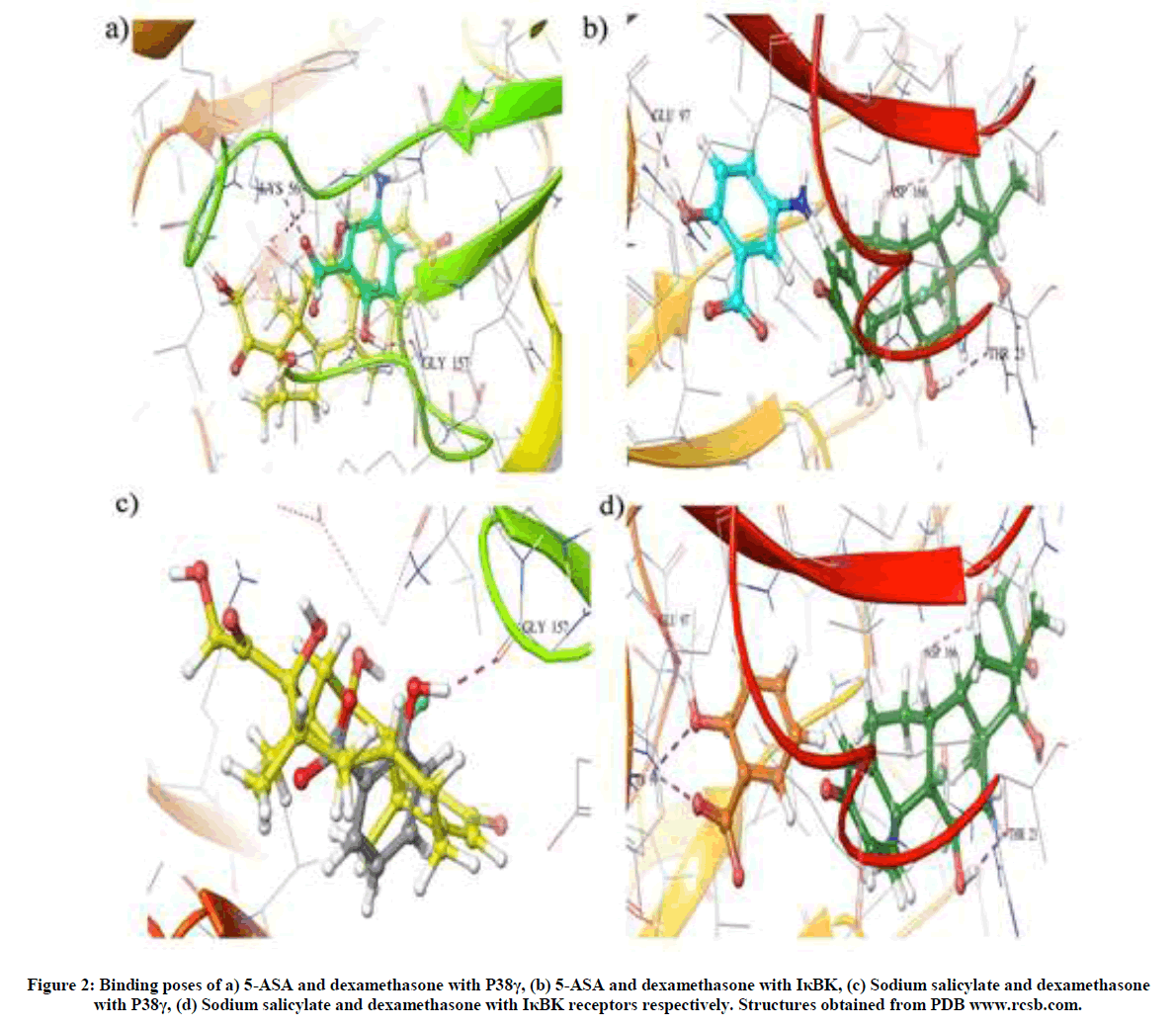
Figure 2: Binding poses of a) 5-ASA and dexamethasone with P38γ, (b) 5-ASA and dexamethasone with IκBK, (c) Sodium salicylate and dexamethasone with P38γ, (d) Sodium salicylate and dexamethasone with IκBK receptors respectively. Structures obtained from PDB www.rcsb.com.
| Ligand code | 5-Aminosalicylic acid | Sodium salicylate | Dexamethasone |
|---|---|---|---|
| P-38γ | -8.6001 | -8.3262 | -8.4042 |
| IκB Kinase | -6.4006 | -6.1475 | -6.2062 |
Table 1: Docking scores of 5-ASA, sodium salicylate and dexamethasone with P38γ, IκBK and JNK pathway
Effect of 5-ASA and sodium salicylate on in vitro cytotoxicity
To determine the cytotoxicity of 5 ASA and SS, RAW 264.7 monocyte-macrophage cells were incubated with the test drugs at 5 different concentrations overnight and were subject to UV absorbance for optical density. 5-ASA at a dose of 36.4 μg/ml has shown cytotoxicity in half of the plated cell population, while SS revealed the same at a dose of 43.7 μg/ml. In the case of the standard (Dexamethasone) the CTC50 was observed at a dose of 29.6 μg/ml respectively (Table 2).
| S. No. | Name of the compound | IC50 mg/ml |
|---|---|---|
| 1. | 5-aminosalicylic acid | 36.4 |
| 2. | Sodium salicylate | 43.7 |
| 3. | Dexamethasone | 29.6 |
Table 2: IC50 values of 5-ASA, sodium salicylate and dexamethasone in RAW 264.7 cell lines using MTT assay
Potential of 5-ASA and Sodium Salicylate to induce apoptosis in RAW 264.7 cell lines
The apoptotic study was performed with both the drugs as they inhibit NFκB. NFκB plays a pivotal role in inducing apoptosis. 5-ASA induced total apoptosis in plated cells at a concentration of 73.5 μg/ml, while there was no induction of apoptosis at a concentration of 18.3 μg/ml. Similarly SS induced total apoptosis in plated cells at a concentration of 87.3 μg/ml, while no apoptosis was observed in case of 21.8 μg/ml concentration respectively (Figure 3).
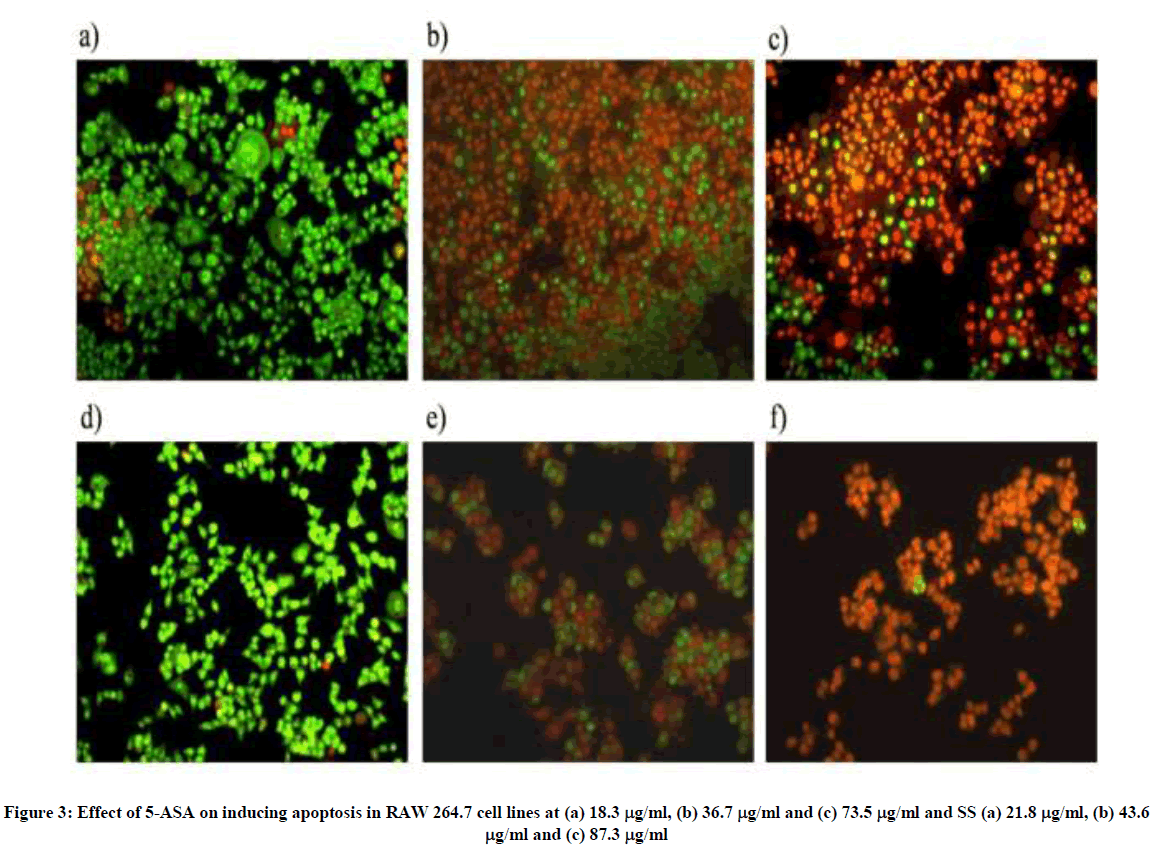
Figure 3: Effect of 5-ASA on inducing apoptosis in RAW 264.7 cell lines at (a) 18.3 μg/ml, (b) 36.7 μg/ml and (c) 73.5 μg/ml and SS (a) 21.8 μg/ml, (b) 43.6 μg/ml and (c) 87.3 μg/ml
Effect of 5-ASA and SS on IκB kinase and P38-γ expression
5-ASA and SS were subjected to mechanistic evaluation by determining the protein expression. In cells treated with LPS the western blot analysis revealed a significant increase in the expression of IκB kinase in RAW cells which was significantly reversed in cells treated with 5- ASA and SS. Similar findings were observed in case of P38-γ expression except for the highest tested dose level of SS (Figure 4).
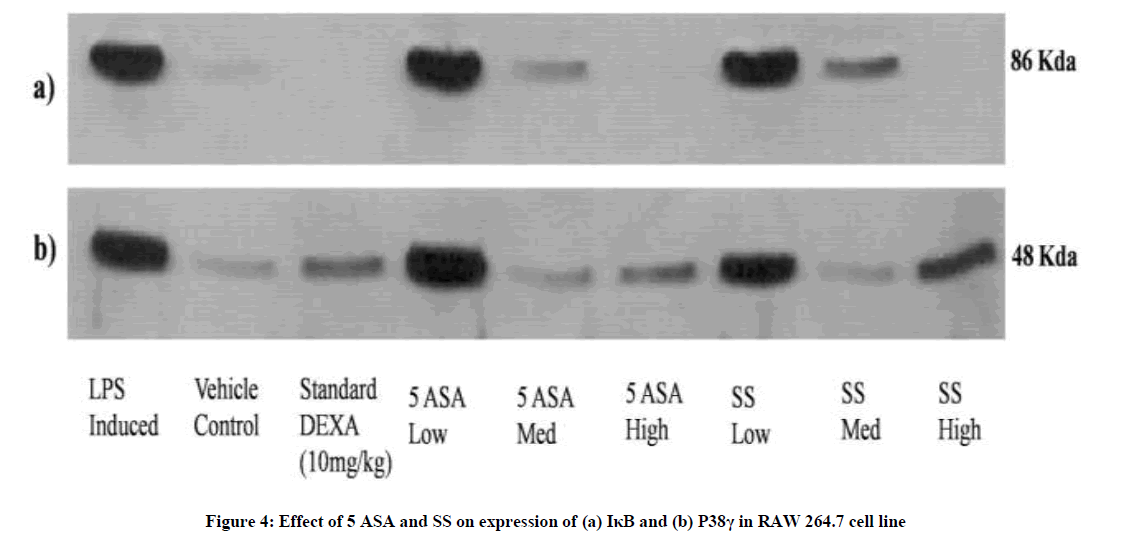
Figure 4: Effect of 5 ASA and SS on expression of (a) IκB and (b) P38γ in RAW 264.7 cell line
Effect of 5-ASA and sodium salicylate on levels of TNF-alpha, IL-6 and IL-13 in RAW 264.7 cells
TNF-α is a TH2 cytokine involved in aggravating allergic reactions by initiating TNF receptor mediated activation of NFκB. Similarly, IL-6 and IL-13 are also TH2 cytokines involved in recruiting eosinophils and activating B-cells to produce IgE. Both the drugs were evaluated for their potential to regulate the levels of the aforementioned cytokines in LPS stimulated RAW 264.7 cells. LPS induced cells have significantly increased the levels of cytokines compared to the vehicle control (15.86 ± 6.12 to 148.1 ± 15.38 in case of TNF-α, 20.10 ± 7.06 to 123.2 ± 12.71 for IL-6 and 22.44 ± 8.90 to 70.05 ± 17.54 for IL-13). Cells treated with 5-ASA have shown a significant dose dependent decrease in the levels of TNF-α, IL-6 and IL-13 compared to LPS control except for the lowest test dose which has no significant activity (Figure 5). In case of Sodium salicylate treated cells, potent activity was seen in the regulation of TNF-α and IL-6 at all tested levels while only the highest tested dose level has shown significant activity in regulating the levels of IL-13.
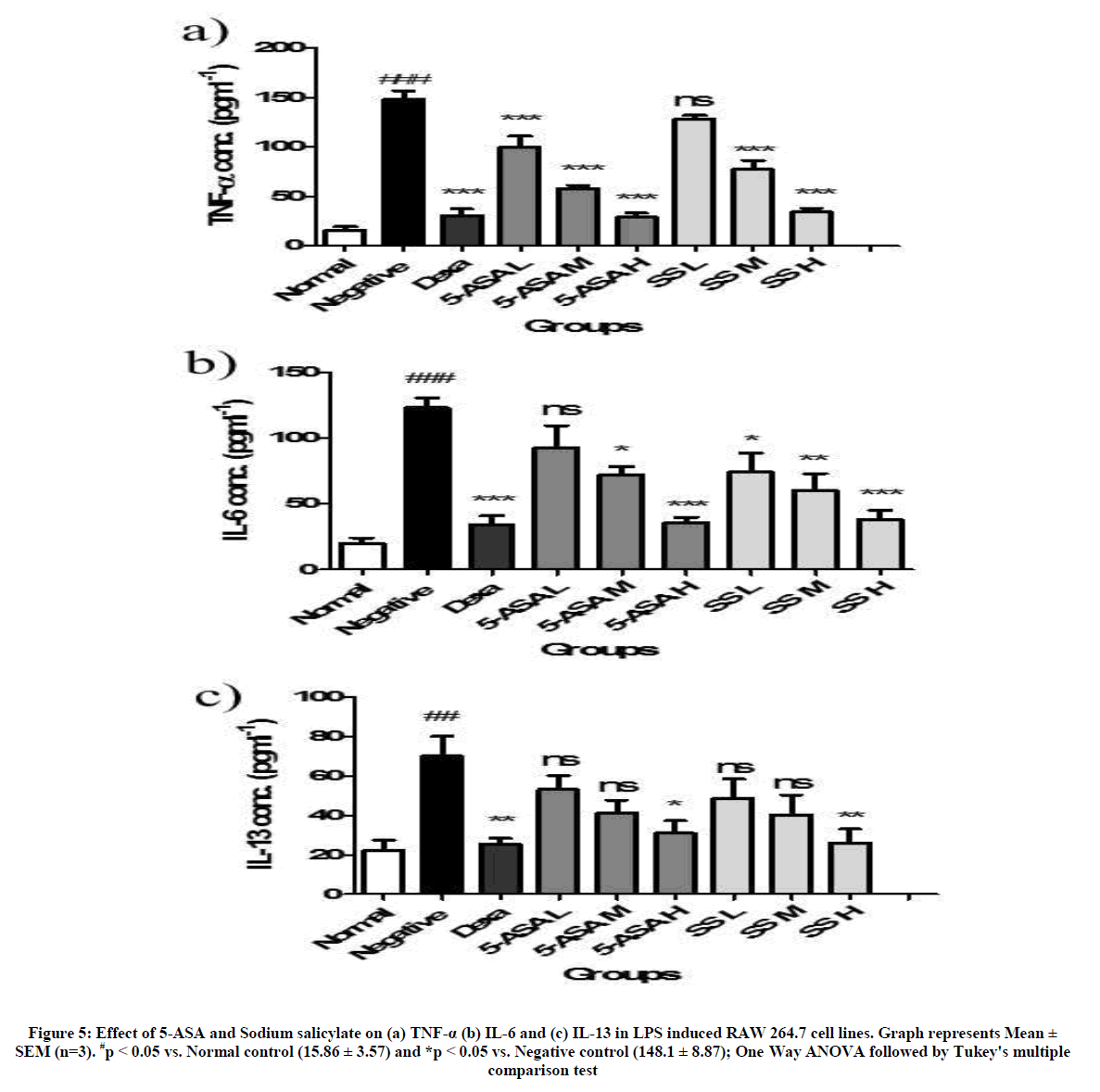
Figure 5: Effect of 5-ASA and Sodium salicylate on (a) TNF-α (b) IL-6 and (c) IL-13 in LPS induced RAW 264.7 cell lines. Graph represents Mean ± SEM (n=3). #p < 0.05 vs. Normal control (15.86 ± 3.57) and *p < 0.05 vs. Negative control (148.1 ± 8.87); One Way ANOVA followed by Tukey's multiple comparison test
Effect of 5-ASA and sodium salicylate on nitrate and nitrite levels
The levels of nitrate and nitrite are proportionate to the level of nitric oxide synthase released during oxidative stress in a cell. Cells induced with LPS alone have shown a significant increase in the levels of nitrate and nitrate proportionately (64.94 ± 16.07 to 196.20 ± 33.89 and 28.21 ± 14.10 to 63.08 ± 17.55 respectively). This was significantly decreased in case of cells treated with 5-ASA and sodium salicylate in a dose dependent manner except for the lowest tested dose level which was not significant (Figure 6).
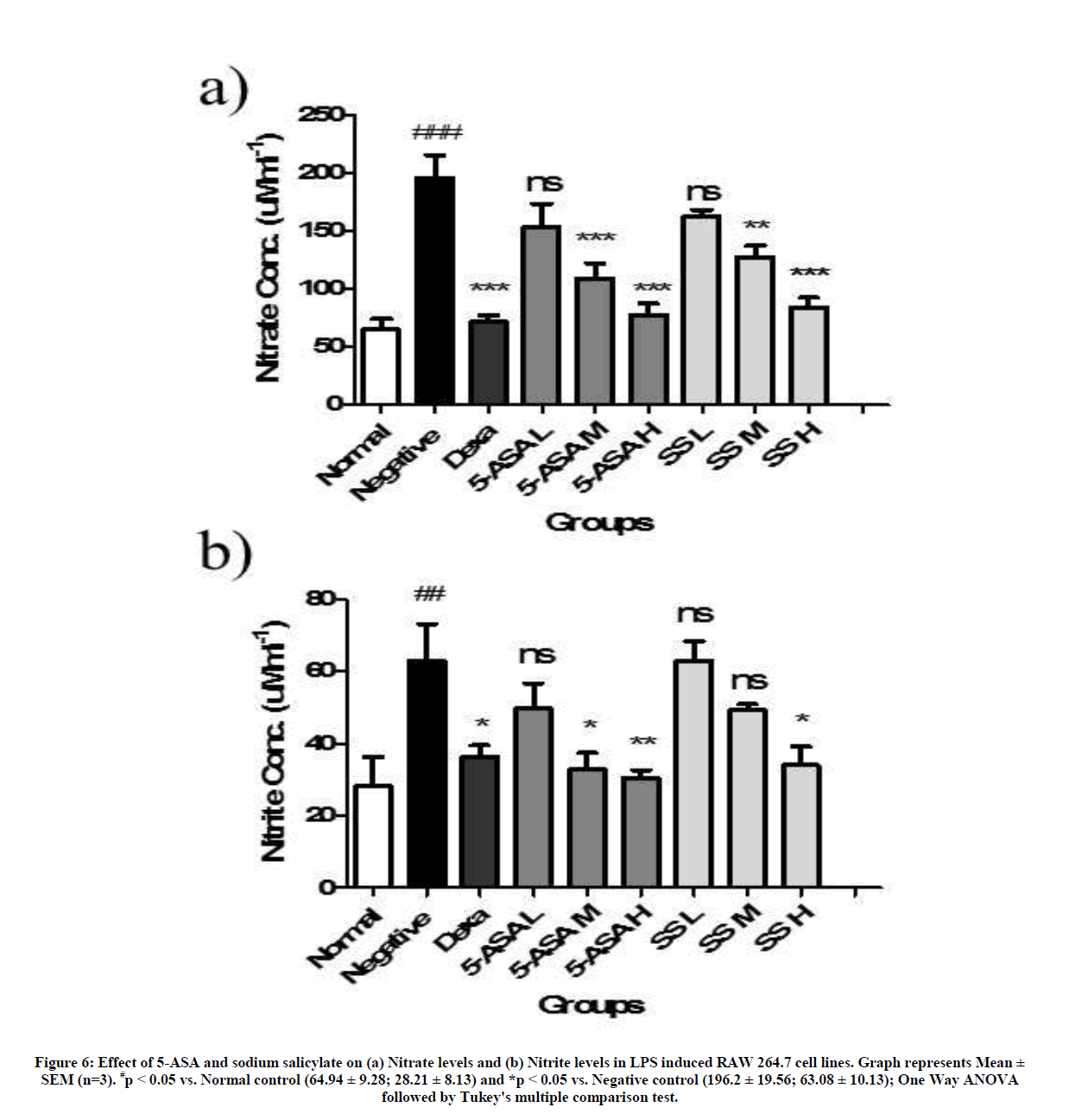
Figure 6: Effect of 5-ASA and sodium salicylate on (a) Nitrate levels and (b) Nitrite levels in LPS induced RAW 264.7 cell lines. Graph represents Mean ± SEM (n=3). #p < 0.05 vs. Normal control (64.94 ± 9.28; 28.21 ± 8.13) and *p < 0.05 vs. Negative control (196.2 ± 19.56; 63.08 ± 10.13); One Way ANOVA followed by Tukey's multiple comparison test.
Effect of 5-ASA and sodium salicylate on myeloperoxidase and malondialdehyde levels
MDA is estimated to quantify the degree of lipid peroxidation in lungs while MPO is estimated for at what extent of cell injury and denaturation of proteins. RAW 264.7 Cells induced with LPS alone have shown a significant increase in the levels of MPO and MDA proportionately (0.145 ± 0.023 and 63.99 ± 3.73 respectively) when compared with normal group (0.034 ± 0.0036; 22.55 ± 4.90). This was significantly decreased in case of cells treated with 5-ASA and sodium salicylate in a dose dependent manner except for the lowest tested dose level which was not significant (Figure 7).
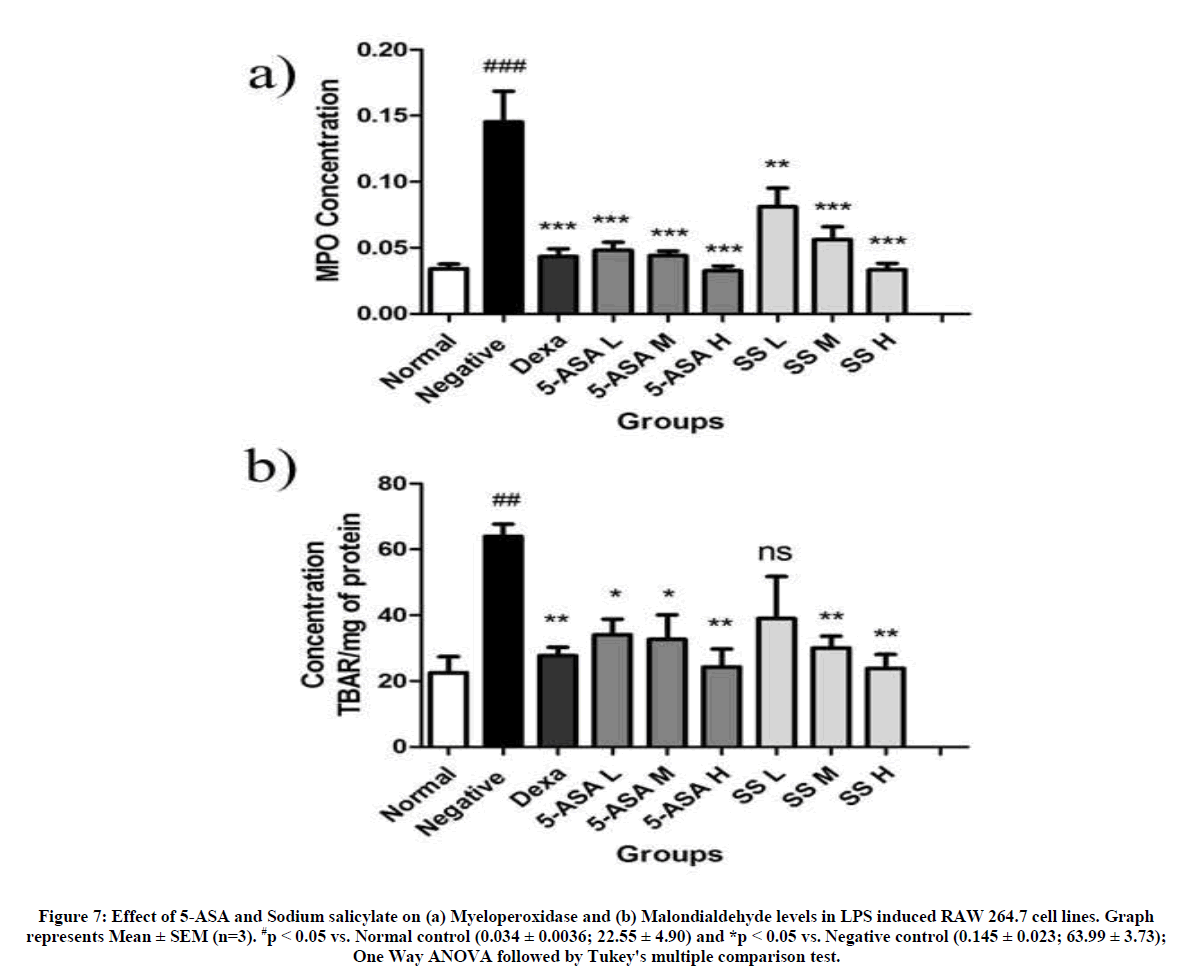
Figure 7: Effect of 5-ASA and Sodium salicylate on (a) Myeloperoxidase and (b) Malondialdehyde levels in LPS induced RAW 264.7 cell lines. Graph represents Mean ± SEM (n=3). #p < 0.05 vs. Normal control (0.034 ± 0.0036; 22.55 ± 4.90) and *p < 0.05 vs. Negative control (0.145 ± 0.023; 63.99 ± 3.73); One Way ANOVA followed by Tukey's multiple comparison test.
Discussion
In this study, an in vitro evaluation of 5-ASA and sodium salicylate as potential candidates for their immunosuppressive activity in allergic asthma has been presented. Nuclear factor κB and P38 MAP kinases play a crucial in regulating the immune responses of a cell during allergic asthma. The in silico findings reveal strong binding affinity of 5-ASA and Sodium Salicylate with IκBK and P38γ, suggesting their ability to inhibit the two proteins. In vitro cytotoxicity study of 5-ASA and sodium salicylate in RAW 264.7 cells had established IC50 values of 36.4 ± 4.67 and 43.7 ± 8.17 μg/ml respectively while dexamethasone as standard drug revealed an IC50 value of 29.6 ± 6.49 μg/ml. The Apoptotic study further verified the innocuous nature of the drug candidates at 18.3 and 21.8 μg/ml respectively for 5-ASA and sodium salicylate.
Followed by the determination of safety profile, the two drug candidates 5 ASA and sodium salicylate were subjected to mechanistic evaluation by determining the protein expression using western blot analysis. NFκB, being a pleiotropic transcription factor, plays a predominant role in allergic asthma by up-regulating the expression of cytokines, chemokines, adhesion molecules and inflammatory enzymes [2]. NFκB is located in the cytosol in a dormant state bound to IκB. IκBK phosphorylates IκB resulting in the activation of NFκB [3]. The western blot analysis has revealed significant increase in the expression of IκBK in LPS induced RAW 264.7 cells which was significantly reversed in cells treated with 5- ASA and Sodium salicylate at test doses in a dose dependent manner. Both 5-ASA and sodium salicylate have significantly suppressed the expression of IκBK at the highest tested dose of 30 μg/ml compared to LPS control. P 38γ, a subgroup of P38MAP kinases dwells in the cytosol which is activated by stress or mitogenic stimuli by many upstream signals which may be P38 specific (MKK3/6), MAPKK independent signals (TAB1) and MAPKK’s (MKK4) [36]. The activated p 38γ results in the activation of MAP kinase-activated protein kinase 2 (MAPK 2 or MK2) [37]. MK2 and MK3 are evidenced to activate heat shock protein 27(HSP27) [38], Lymphocyte-Specific Protein 1 (LSP1) [39], cAMP response element-binding protein (CREB) [40], transcription factor ATF1 [41], SRF [42] and tyrosine hydroxylase [43]. Apart from this it is also documented to indirectly influence Activator Protein 1 (AP-1) which is a key transcription factor involved in the pathophysiology of allergic asthma. P38 MAPK is involved in the transcriptional regulation of cytokines, chemokines, adhesion molecules and inflammatory enzymes. Both 5-ASA and sodium salicylate have significantly decreased the expression of P38γ in a dose dependent manner except for the highest tested dose (30 μg/ml) of Sodium salicylate. This might be due the interaction of sodium salicylate at higher doses with upstream proteins of P38 MAP kinases like caspases [45,46]. The levels of TNF-α, IL-6 and IL-13 have been regulated by 5 ASA and sodium salicylate at tested dose levels dose dependently when compared to LPS control which correlates with western blot analysis. Similarly, nitrate, nitrite, malondialdehyde and myeloperoxidase levels were regulated to normal when at all three tested dose levels of 5-ASA and sodium salicylate dose dependently.
Conclusion
The findings from the present study demonstrate the potential of 5-ASA and sodium salicylate as NFκB and P38 MAPK inhibitors in vitro. The results clearly indicate the effect of 5-ASA and sodium salicylate on regulating the expression of cytokines, nitrate and nitrite levels, malondialdehyde and myeloperoxidase by inhibiting the activation of NFκB and P38γ MAP kinase, suggesting the two drug candidates as lead molecules for allergic asthma.
Acknowledgement
The authors would like to acknowledge Late Dr. M.N. Sathish Kumar for his immense support during the tenure of the study. The author Mr. K. Rama Satyanarayana Raju would like to thank Indian Council of Medical research for providing financial support in the form of senior research fellowship (Grant No.: 45/01/2014-NAN/BMS). The authors would also like to thank JSS University, Mysore and JSS College of Pharmacy for providing the infrastructure to carry out the study.
References
- T. Phillips, L. Hoopes, Nature Education., 2008, 1(1), 119.
- P.J. Barnes, M. Karin, N. Engl. J. Med., 1997, 336(15), 1066-1071.
- J.C. Kips, J.H. Tavernier, G.F. Joos, R.A. Peleman, R.A. Pauwels, Clin. Exp. Allergy., 1993, 23(4), 247-250.
- J. Rouse, P. Cohen, S. Trigon, M. Morange, A. Alonso-Llamazares, D. Zamanillo, T. Hunt, A.R. Nebreda, Cell., 1994, 78(6), 1027-1037.
- T. Zarubin, J. Han, Cell Res., 2005, 15(1), 11-18.
- G.V. Johnson, C.D. Bailey, Exp. Neurol., 2003, 183(2), 263-68.
- E. Hollenbach, M. Neumann, M. Vieth, A. Roessner, P. Malfertheiner, M. Naumann, FASEB J.,2004, 18(13), 1550-1552.
- D.G. Perregaux, D. Dean, M. Cronan, P. Connelly, C.A. Gabel, Mol. Pharmacol., 1995, 48(3), 433-442.
- Y. Jiang, C. Chen, Z. Li, W. Guo, A.J. Gegner, S. Lin, J. Han, J. Biol. Chem., 1996, 271(30), 17920-17926.
- C. Lechner, M.A. Zahalka, J.F. Giot, M.P.H. Moler, A. Ullrich, Proc. Natl. Acad. Sci. USA., 1996, 93(9), 4355-4359.
- Z. Li, Y. Jiang, R.J, Ulevitch, J. Han, Biochem. Biophys. Res. Commun., 1996, 228(2), 334-340.
- Y. Jiang, H. Gram, M. Zhao, L. New, J. Gu, L. Feng, F. Di Padova, R.J. Ulevitch, J. Han, J. Biol. Chem., 1997, 272(48), 30122-30128.
- S. Kumar, P.C. McDonnell, R.J. Gum, A.T. Hand, J.C. Lee, P.R. Young, Biochem. Biophys. Res. Commun., 1997, 235(3), 533-538.
- S.K. Hanks, T. Hunter, FASEB J., 1995, 9(8), 576-596.
- F. Yan, B.D. Polk, J. Biol. Chem., 1999, 274(51), 36631-36636.
- K. Kiss, J. Kiss, E. Rudolf, M. Cervinka, J. Szeberenyi, J. Biochem. Biophys. Methods., 2004, 61(1-2), 229-240.
- K.R. Raju, M.N. Kumar, S. Gupta, S.T. Naga, J.K. Shankar, V. Murthy, S.R. Madhunapanthula, S. Mulukutla, N.S. Ambhore, S. Tummala, V.J. Vishnuvarthan, A. Azam, K. Elango, Pulm. Pharmacol. Ther., 2014, 29(2), 209-216.
- D. Shivakumar, J. Williams, Y. Wu, W. Damm, J. Shelley, W. Sherman, J. Chem. Theory Comput., 2010, 6(5), 1509-1519.
- S. Liu, Y.R. Misquitta, A. Olland, M.A. Johnson, K.S. Kelleher, R. Kriz, L.L. Lin, M. Stahl, L. Mosyak, J. Biol. Chem., 2013, 288(30), 22758-22767.
- S. Bellon, M.J. Fitzgibbon, T. Fox, H.M. Hsiao, K.P. Wilson, Structure., 1999, 7(9), 1057-1065.
- G.M. Sastry, M. Adzhigirey, T. Day, R. Annabhimoju, W. Sherman, J. Comput. Aided Mol. Des., 2013, 27(3), 221-234.
- J.R. Greenwood, D. Calkins, A.P. Sullivan, J.C. Shelley, J. Comput. Aided Mol. Des., 2010, 24(6-7), 591-604.
- V. Reetu, Kumar, Int. J. Pharm. Sci. Res., 2012, 3(3), 805-810.
- G.M. Sastry, S.L. Dixon, W. Sherman, J. Chem. Inf. Model., 2011, 51(10), 2455-2466.
- T. Mosmann, J. Immunol. Methods., 1983, 65(1-2), 55-63.
- J. Hambleton, S.L. Weinstein, L. Lem, A.L. Defranco, Proc. Natl. Acad. Sci. USA., 1996, 93(7), 2774-78.
- K. Liu, P.C. Liu, R. Liu, X. Wu, Med. Sci. Monit. Basic Res., 2015, 21, 15-20.
- M.M. Bradford, Anal. Biochem., 1976, 72, 248-254.
- T.J. Toward, K.J. Broadley, Br. J. Pharmacol., 2000, 131(2), 271-281.
- O.I. Aruoma, B. Halliwell, M.J. Laughton, G.J. Quinlan, J.M. Gutteridge, Biochem. J., 1989, 258(2), 617-620.
- P.P. Bradley, R.D. Christensen, G. Rothstein, Blood.,1982, 60(3), 618-622.
- World asthma days. Global burden of asthma report, Embargoed, 2016.
- http://www.who.org/.
- www.sancd.org/uploads/pdf/Asthma_factsheet.pdf.
- S. Ganesh Kumar, K.C. Premarajan, S. Sarkar, K. Sahu Swaroop, S. Ambika, A. Antony, Curr. Pediatr. Res., 2012, 16(2), 159-163.
- T. Zarubin, J. Han, Cell Res., 2005, 15(1), 11-18.
- M.M. McLaughlin, S. Kumar, P.C. McDonnell, S.V. Horn, C.J. Lee, P.G. Livi, R.P. Young, J. Biol. Chem., 1996, 271(14), 8488-8492.
- D. Stokoe, K. Engel, D.G. Campbell, P. Cohen, M. Gaestel, FEBS Lett., 1992, 313(3), 307-313.
- C.K. Huang, L. Zhan, Y. Ai, J. Jongstra, J. Biol. Chem.., 1997, 272(1), 17-19.
- Y. Tan, J. Rouse, A. Zhang, S. Cariati, P. Cohen, M.J. Comb, EMBO J., 1996, 15(17), 4629-4642.
- O. Heidenreich, A. Neininger, G. Schratt, R. Zinck, M.A. Cahill, K. Engel, A. Kotlyarov, R. Kraft, S. Kostka, M. Gaestel, A. Nordheim, J. Biol. Chem., 1999, 274(20), 14434-1443.
- G. Thomas, J. Haavik, P. Cohen, Eur. J. Biochem., 1997, 247(3), 1180-1189.
- K.R. Mahtani, M. Brook, J.L. Dean, G. Sully, J. Saklatvala, A.R. Clark, Mol. Cell Biol., 2001, 21(19), 6461-6469.
- A.J. Waskiewicz, A. Flynn, C.G. Proud, J.A. Cooper, EMBO. J., 1997, 16(8), 1909-1920.
- P.A. Henkart, Immunity., 1996, 4(3), 195-201.
- S. Huang, Y. Jiang, Z. Li, E. Nishida, P. Mathias, S. Lin, R.J. Ulevitch, G.R. Nemerow, J. Han, Immunity., 1997, 6(6), 739-749.